Nutrición pre, intra y post competencia 4º Parte: Los lípidos en el metabolismo del movimiento Dra. Patricia Minuchin.
La función de las grasas es la de proveer energía, pero tiene el inconveniente del tiempo que tarda en oxidarse. La oxidación de las grasas no puede soportar una intensidad de entrenamiento mayor al 60% de la potencia aeróbica máxima.
Tipos de lípidos: 1) Triglicéridos(formados por glicerol y tres AG) formadas por AG saturados, monosaturados y poliinsaturados. Son el 98% de las grasas que ingresan por nuestra dieta, y el 90% de los lípidos presentes en nuestro cuerpo. Su función es energética. 2) Fosfolípidos (formados por glicerol, dos AG y un grupo fosforado) ej: lecitina, esfingomielina, cardiolipinas. Según el tipo de AG que tenga (polinsaturado o saturado le dará a la membrana que conforme, distintas propiedades. Los polinsaturado (pescados, aceites de semillas) hacen más permeable a la membrana al O2. 3) Colesterol (precursor de hormonas, vitamina D, ácidos biliares) Los AG esenciales son el LINOLEICO Y LINOLÉNICO ( de origen vegetal y en algunos pescados). Aparte de los beneficios biomecánicos y térmicos, las grasas: 1)Tienen mas del doble de Kcal/g Las grasas contienen 9 Kcal/g, a diferencia de los HC: 4 Kcal/g. 2) No se depositan con agua (mas eficiencia por unidad de peso) Un gramo de glucógeno fija 2.7 ml de agua. 3) sus depósitos no son tan limitados como los HC (hígado: 100 g)(músculo: 200-600 g) ( Plasma: 20 g). Entre 300 y 700 gs= 1200 y 2800 kcal en total. 4) se sintetizan los eicosanoides.(De los Fosfolípidos de la membrana)Provienen especialmente de los aceites de pescado (omega 3). La literatura española (Javier Gallego y Vicente) promueve la ingesta del aceite de oliva debido a su gran contenido de monoinsaturados que aumentan los FL de membrana menos oxidables (con la disminución de radicales libres) y mayor producción de eicosanoides. La oxidación se produciría sobre los dobles enlaces de los Carbonos (abundantes en los poliinsaturados)
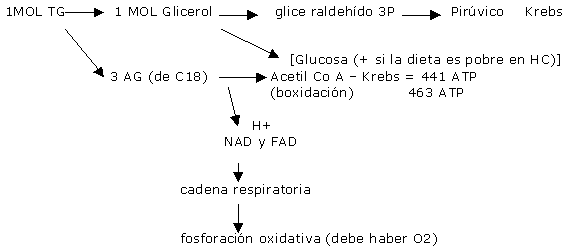
La contra de las grasas: 1) tardan mas tiempo en oxidarse. 2) solo pueden ser usados por vía aeróbica. 3) hay tejidos que usan exclusivamente glúcidos (SNC) Para oxidar una molécula de glucosa (6 carbonos) se obtienen 38 ATP y se usan 6 moléculas de O2. Y para oxidar una molécula de un AG (esteárico) se obtienen 147 ATP y requiere 26 moléculas de O2. (por lo que se necesita mas O2 para degradar los AG que para la glucosa aún para producir la misma cantidad de ATP) Los HC y la grasa son la fuente principal de combustible durante AF de resistencia. Y la capacidad de oxidar grasa es parte del entrenamiento invisible.
Aquel deportista que pueda activar más rápido las grasas en un trabajo de resistencia, a su vez ahorrara glucógeno, con lo que se retrasará la producción de ácido láctico y la fatiga. Cociente Respiratorio (CR) indica que combustible se está utilizando. CR= CO2 (aire espirado)/ O2 (consumido) Si la dieta es rica en grasas el CR=0.7 Si la dieta es rica en HC el CR= 0.95 Sin embargo cuando la dieta es rica en grasas la percepción de la fatiga es antes. Aparte no se debe olvidar que las grasas no son saludables cuando se consumen en exceso. La mayor cantidad de grasa se encuentra alojada en el tejido adiposo subcutáneo y visceral. El exceso de AG, se convertirá en TG que se depositará en las células grasas del tejido adiposo. Pero en menor grado también se deposita dentro de las células musculares como vacuolas. Este pool es de 300 grs aprox. Y las fibras tipo I tienen mayor cantidad de TG que las tipo II.
Las reservas grasas en el organismo son:
1) AGL plasmáticos. 2) TG de las VLDL y QM plasmáticos (LPP ó lipoproteínas plasmáticas). 3) TG intramusculares 4) TG del tejido adiposo
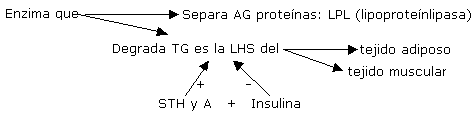
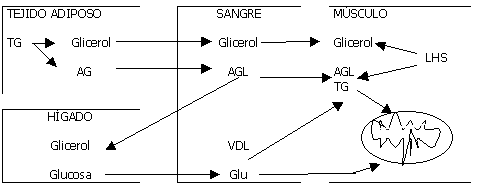
1) AGL: Son el resultado de la lipólisis de los TG del tejido adiposo, muscular y en menor grado de la lipoproteínas plasmáticas.( Se usan especialmente cuando la intensidad del ejercicio es del 25 %). Las células musculares también poseen LHS (lipasa hormono sensible) que es la enzima que hidroliza los TG en glicerol y AG..
2) Otra fuente posible de AG, son los TG de las LP (lipoproteínas). La LPL (lipoproteín-lipasa) de la pared vascular hidroliza algunos de los TG de las LP (aunque en realidad solo aportan el 5% de la energía derivada de los AG)La actividad de la LPL aumenta significativamente luego del entrenamiento y luego de un dieta rica en grasas, sin embargo la contribución de los TG de las LP plasmáticas es mínima durante la AF.
3)TG intramuscular (declina el uso de AGL plasmáticos y aumenta el de los Tgim cuando la I del ej es del 60% del Vo2). Las fibras tipo I contienen mas TG im que las tipo II. Son otra fuente de energía que se encuentra cerca de las mitocondrias, y disminuyen significativamente postejercicio.Los AG provenientes de los TG im pueden ser oxidados en el mismo músculo o ser liberados a la sangre.
4)AG del tejido adiposo.
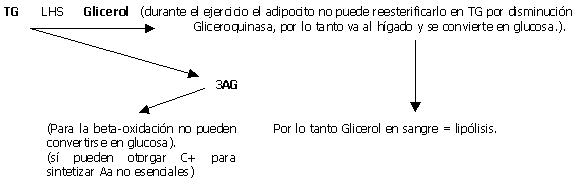
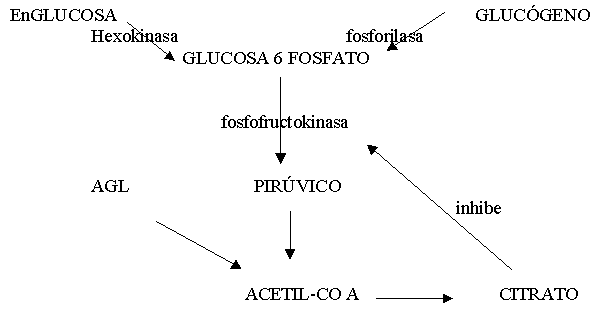
Una vez en el músculo los AGL unidos a la coenzima A , se transforman en Acil-coA , la carnitina lo transporta por la membrana mitocondrial donde es se oxida directamente o a traves del ciclo de Krebs. Se cree que los AG de cadena corta y media pueden difundir a través de la membrana mitocondrial por medio de otros carriers para finalmente ser B-oxidados. El producto de degradación del catabolismo de AG son los cuerpos cetónicos(aceto-acetato y B-OH-butírico). Estos pueden ser utilizados como energía para la conversión de glicerol en glucosa. Antes de ser oxidados deben ser transportados. La LHS (lipasa hormonosensible) que a su vez es estimulada por la adrenalina e inhibida por la insulina, estimula la movilización de AG del tejido adiposo y los del tejido muscular. y se divide en glicerol y 3 AG. Por lo que la aparición de glicerol en sangre es considerado como un valor de la tasa de lipólisis. (El glicerol del adipocito no puede ser reesterificado en AG con rapidez debido al bajo nivel de Gliceroquinasa). Los AGL pueden reesterificarse en TG (suprimido por el ejercicio), o ser transportados por la albúmina a los tejidos para ser degradados como energía.
En reposo, el 70% de los AGL se reesterifican en TG. El ejercicio disminuye la
reesterificación y aumenta la lipólisis, con lo que aumentan los
AGL.
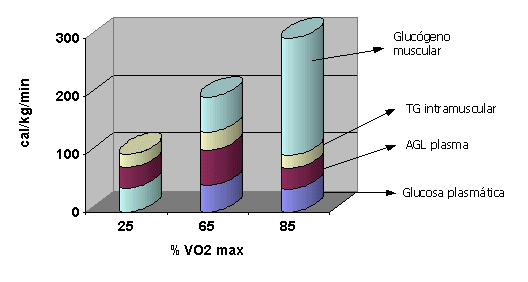
El entrenamiento produce mayor utilización de AG debido a: 1)Aumento de enzimas oxidativas (son 13 para los AG). 2)Aumento del número de mitocondrias. 3)Incremento de la utilización de AG intramusculares. 4)Aumento de la movilización de AG desde el tejido adiposo (adrenalina) 5)Aumento de la captación muscular de AG En reposo el 70% de los AGL del tejido adiposo es reesterificado, pero durante el ejercicio son transportados por la albúmina donde se dirigen al músculo para ser utilizados allí. A una intensidad baja (25% del Vo2 max.) la mayor parte de la grasa oxidada proviene de los AG L plasmáticos . A una intensidad del 65% disminuye el aporte de los plasmáticos y aumenta el del tejido intramuscular (que pueden aportar hasta el 50% de los AGL para oxidación). A una intensidad del 85% disminuye la oxidación proveniente de los AG plasmáticos y de los TG intramusculares para aumentar la oxidación del glucógeno muscular. Cuando la intensidad del ejercicio supera el 60% la utilización de grasas declina para aumentar el uso de HC.En el siguiente gráfico Romijn y colaboradores nos muestran la contribución de los cuatro sustratos principales energéticos, luego de 30 minutos de ejercicio a distintas intensidades con ayuno previo de la noche anterior.
El entrenamiento acelera la utilización de Tgim respecto de los AGL plasmáticos. Durante el ejercicio: Durante los primeros 15 minutos de ejercicio, las concentraciones de AGL disminuye pues la captación muscular excede la liberación por lipólisis. A partir de allí la lipólisis hace que comiencen a aumentar nuevamente en plasma camino al músculo. La oxidación aumenta con la duración del ejercicio. Relativamente, la oxidación de las grasas será máxima con intensidades bajas. ( Edwards reportó 1 g/minuto de oxidación a una intensidad del 60%) Christensen y Hansen observaron que el uso de AG puede ser utilizado más (1.5g/min) cuando se consume una dieta rica en grasas. A intensidades del 80% del máximo, la grasa como combustible decrece significativamente.
Ayuno y AF: El ayuno predispone a aumentar la disponibilidad de AG en sangre. Sin embargo un estudio realizado en ciclistas (Lo) probó que si bien la disponibilidad de AG mejora, la performance empeora por el ayuno. Al aumentar la oxidación de AG, aumenta el citrato. El citrato inhibe la PFK, con aumento de la GLU-6 P por lo que se reduce la Glucólisis.
Por otro lado, recordemos que la insulina inhibe a la LHS, por lo que en ejercicios que mantienen la insulina por debajo de los valores de reposo, se supone que ésto libera la inhibición. El ayuno produce por éste mecanismo(disminución de la insulina) la liberación de la LHS, con consiguiente aumento de la lipólisis. A su vez no debemos confundir lipólisis con oxidación de AG.
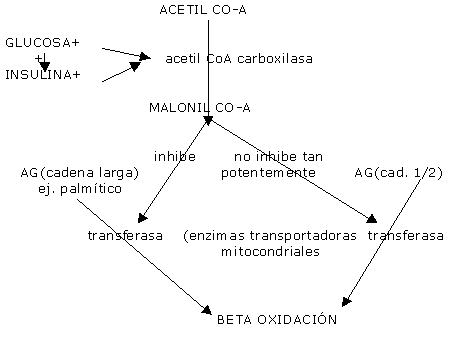
Factores que regulan la lipólisis: Aparte de las catecolaminas (más la adrenalina que la noradrenalina) que estimulan a la LHS del tejido adiposo y del intramuscular, existirían factores intracelulares que regulan la lipólisis. Si le damos glucosa previa al ejercicio, la oxidación disminuye y la lipólisis también. Aparentemente la ingesta de HC parece limitar la entrada de AG a nivel mitocondrial. Horowitz demostró que si la lipólisis disminuye, disminuye la oxidación de AGL. Pues al inyectar TG y heparina (estimuladora de la LPL) la oxidación aumentaba, aún dando glucosa previa. Aunque no tanto como en el ayuno. Cuando la lipólisis excede la oxidación de AG, el factor limitante podría ser la entrada de AG a la mitocondria y ésta última podría ser causado por el mayor flujo glucolítico. Se ha probado la suplementación de AG de cadena larga (12 o mas carbonos) ( que sabemos son absorbidos como QM por vía linfática y tardan 3-4 hs para llegar a la circulación general Es decir que no son útiles para la pre e intracompetencia,, aparte demoran el vaciado gástrico. Los TG de cadena mediana (6 a 10 carbonos)y corta, son absorbidos y aparecen en sangre mas rápidamente , debido a que por vía portal se dirigen al hígado y de allí a la sangre general .Son más solubles en agua y a t°C ambiente son fluidos por lo que no retrasan el vaciamiento gástrico. Por lo que se ha sugerido su suplementación (30 gs), pero no tuvieron efectos positivos en la performance, ni siquiera ingeridos con HC. Posiblemente sea por el mayor flujo glucolítico (aparte solo pueden ser ingeridos en pequeñas cantidades para no causar malestar gastrointestinal). Los AGL solo podrían entrar a la célula muscular vía difusión facilitada por proteínas(barrera limitante para el uso de AG) Coyle concluye que la oxidación de AG está aumentada al aumentar la densidad mitocondrial. Y el número de mitocondrias aumenta con el entrenamiento de la resistencia (aún si la intensidad del esfuerzo es del 70% del Vo2 máximo)
El mecanismo sería el siguiente:
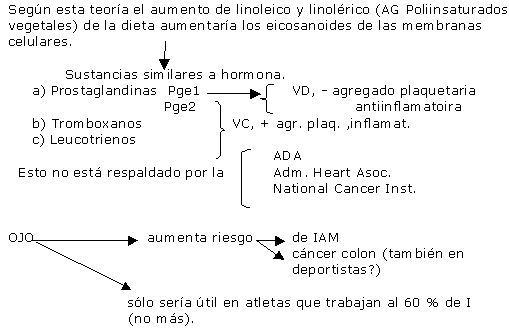
Es decir que la glucosa y la insulina limitan el transporte mitocondrial de los AG de cadena larga pero no tanto a los de cadena mediana, debido a una inhibición directa de su transportador mitocondrial y no por un fenómeno pasivo de flujo glucolítico. Todo ésto minimiza la aparición de AG plasmáticos, cuando no pueden ser oxidados por el músculo. Existe una dieta promocionada por Barry Sears, que propone un alto % graso y proteico y menos de HC. El propone HC:40% P:30% G:30% Refiere que elevan los eicosanoides , y las prostaglandinas que regulan el sistema hormonal . Los eicosanoides son sustancias derivadas del linoleico y linolénico (AG vegetales esenciales ricos en poliinsaturados). Son sustancias hormonosímiles llamadas: 1)Prostaglandinas : PG E1(vasodilatadora, inhibe la agregación plaquetaria y es antiinflamatoria). bueno. Aparte ésto mejora la Vo2 máxima( por vasodilatación). estimula la STH y disminuye la fatiga por efecto antinflamatorio. proviene de los aceites de pescado(omega 6). PG E2 (vasoconstrictora, estimula la agregación plaquetaria y es antiinflamatoria). malo deriva del araquidónico y del eicosapentanoico. 2)Tromboxanos (idem PG E2). 3)Leucotrienos.(idem PG E2) Según Barry Sears, la relación porcentual entre los HC y P provocan un balance entre insulina – glucagon que aumentarían los eicosanoides buenos y disminuirían los malos. El balance positivo hacia la insulina aumentaría los malos, que interfieren con el uso de los AG, y disminuyen el VO2. Supuestamente las proteínas y las grasas aumentan el glucagon. Esto no esta respaldado por la ADA, ni la Am. Heart Association, ni el National Cancer Institute . Muy por el contrario en la revista Metabolism en el 96’encontro que los niveles de insulina eran significativamente más bajos en dietas alta en fibras y ricas en HC, y bajas en grasas. Aparte los atletas no mejoran su performance por utilizar mas las grasas, sino por entrenar mejor la cualidad "resistencia". SI BIEN LAS HIPOTESIS DE LA CARGA ALIMENTARIA DE GRASAS MEJORARIAN LA PERFORMANCE, DESDE LA PERSPECTIVA DE SALUD SE RELACIONA con AUMENTAR EL RIESGO DE ENF. CARDIOVASCULARES. Cuando se consume una dieta alta en grasas y baja en HC por 5 días en ciclistas, Kaijser reporta una disminución de la performance, de la resistencia y aumento de la fatiga (por reducción de los depósitos de glucógeno). Sin embargo con dietas que duran mas de 15 días, se produce el aumento de enzimas oxidativas de los AG, si bien en ejercicios al 60% del máximo, aumenta la performance ( Lambert),que es una intensidad que no se usa en la mayoría de los deportes. Pendergast reportó que no hubo cambios del perfil lipídico en corredores de resistencia que consumían dietas desde 17 al 40% del VCT. Pero es sabido que el riesgo es mayor en sedentarios. También es sabido que la dieta rica en grasas genera una insulino resistencia. Sin embargo esto se estudió en obesos, y no en entrenados que poseen mayor aumento de la sensibilidad muscular a la insulina. Guezennec sugiere que un aumento de los AG poliinsaturados provenientes del aceite de pescado, altera los fosfolípidos de la membrana de los glóbulos rojos de tal manera que mejora la difusión del O2. Sin embargo deberá ser mas estudiado para comparar éste efecto con sedentarios.
CONCLUCIONES para grasas: BENEFICIOS Y DESVENTAJAS DE LAS GRASAS:
Una dieta rica en grasas por más de 15 días, podría mejorar la performance, pero deberían estudiarse efectos adversos. Ej. : Barry Scars HC: 40% P : 30% G : 30%
11) Tarnopolski: la mujer oxida + grasas que el varón (en ej. De I moderada 60%). 12) El trabajo intermitente (Essen) moviliza AG más que el L continuo. 13) La carnitina favorece la entrada de AG a la mitocondria (pero el aumento de carnitina no aumenta la gentrada). 14) Guezennec sugiere un aumento de AG PI (aceite de pescado) para que altere la composici{on de los FL de la Membrana de los GR y mejorar la difusión del O2.Sin embargo faltan estudios que corroboren esto. Bibliografía: 1.Berning, J. , Nelson Steen ,S.. Nutrition for Sport and Exercise. N. York: editorial ASPEN. 1998, 2. Delgado Fernandez M., Gutiérrez Saínz A., Castillo Garzón M.J.. Entrenamiento Físico-Deportivo y alimentación. Universidad de Granada: Editorial Paidotribia, 1997; Capítulos 6 3.González Gallego J., Garrido Pastor G., Mataix Verdú J., y col.. Nutrición y ayudas ergogénicas en el deporte. Madrid,: Editorial SINTESIS,1998. 4.Creff Bérard A.F. Dietética Deportiva. Barcelona: editorial Masson. 1985. 5.Wootton Steve. Nutrición y Deporte. España: editorial Acribia.!990. 8. 6.Gonzalez Ruano E. Alimentación del deportista. Madrid: editorial Marbán. 1986. 7.Eduard Coyle. Universidad de texas, Austin.Biochemistry of Exercise. 1998.
|
|
|