![]()
![]()
![]()
3.1 Clasificación
3.2 Anatomía y fisiología
3.3 Crecimiento y reproducción
Los peces generalmente se definen como vertebrados acuáticos, que utilizan branquias para obtener oxígeno del agua y poseen aletas con un número variable de elementos esqueléticos llamados radios (Thurman y Webber, 1984).
Cinco clases de vertebrados poseen especies que pueden ser llamadas peces, pero sólo dos de estos grupos - los peces cartilaginosos (los tiburones y las rayas) y los peces óseos - son generalmente importantes y están ampliamente distribuidos en el ambiente acuático. La relación evolutiva entre los diferentes grupos de peces se muestra en la Figura 3.1.
Los peces son los más numerosos de los vertebrados; existen por lo menos 20.000 especies conocidas y más de la mitad (58 por ciento) se encuentran en el ambiente marino. Son más comunes en las aguas cálidas y templadas de las capas continentales (unas 8.000 especies). En las frías aguas polares se encuentran alrededor de unas 1.100 especies. En el ambiente pelágico del océano, alejado de los efectos terrestres, se encuentran sólo unas 225 especies. Sorprendentemente, en las profundidades de la zona mesopelágica (entre 100 y 1000 metros de profundidad) el número de especies incrementa. Existen unas 1.000 especies de los denominados peces de media agua ("midwater fish") (Thurman y Webber, 1984).
Clasificar todos estos organismos en un sistema no es una tarea fácil, pero el taxonomista agrupa organismos en unidades naturales que reflejan las relaciones evolutivas. La unidad más pequeña es la especie. Cada especie es identificada mediante un nombre científico, constituido por dos partes - el género y el epíteto específico (nomenclatura binomial). El género siempre se escribe con mayúscula y las dos partes siempre van en letras itálicas). Como un ejemplo, el nombre científico del delfín común es Delphinus delphis. El género es una categoría que contiene una o más especies, mientras que el próximo paso en la jerarquía es la familia, que puede contener uno o más géneros. Por lo tanto, el sistema jerárquico total es: Reino: Filo (Phylum): Clase: Orden: Familia: Género: Especie.
El uso de nombres locales o comunes crea generalmente confusión, dado que la misma especie puede tener diferentes nombres en distintas regiones, o por el contrario, el mismo nombre puede estar asignado a diferentes especies, a veces con propiedades tecnológicas diferentes. Por lo tanto, el nombre científico debe ser dado como punto de referencia en cualquier clase de publicación o reporte, la primera vez que la especie sea citada por su nombre común. Para mayor información, se deben consultar: el Consejo Internacional para la Exploración del Mar "Lista de nombres de peces y mariscos" (del inglés International Council for the Exploration of the Sea "List of names of Fish and Shellfish") (ICES, 1966); el Diccionario Políglota de Peces y Productos Pesqueros preparado por la Organización para la Cooperación Económica y el Desarrollo (del inglés Organisation for Economic Cooperation and Development) (OECD, 1990) y el Diccionario Políglota Ilustrado de Animales y Plantas Acuáticas (Comisión de las Comunidades Europeas, 1993).
La clasificación de los peces en cartilaginosos y óseos (los peces no mandibulados son de menor importancia) resulta importante desde el punto de vista práctico y también por el hecho de que estos grupos de peces se deterioran en formas diferentes (sección 5) y varían respecto a su composición química (Sección 4).
Además, los peces pueden ser divididos en especies grasas y especies magras, pero este tipo de clasificación se basa en características biológicas y tecnológicas según se muestra en el Cuadro 3.1.
Cuadro 3.1 Clasificación de los peces
|
Grupo científico |
Características biológicas |
Características tecnológicas |
Ejemplos |
|
Ciclóstomomos |
peces no mandibulados |
|
lampreas, anguilas tiburones, rayas, mantas |
|
Condrictios |
peces cartilaginosos |
alto contenido de urea en el músculo |
|
|
Teleósteos o peces óseos |
peces pelágicos |
pescado graso (lípidos almacenados en el tejido muscular) |
arenque, caballa, sardina, atún |
|
peces demersales |
pescado (blanco) magro, almacena lípidos solamente en el hígado |
bacalao, eglefino, merluza, mero, cherna |
El esqueleto
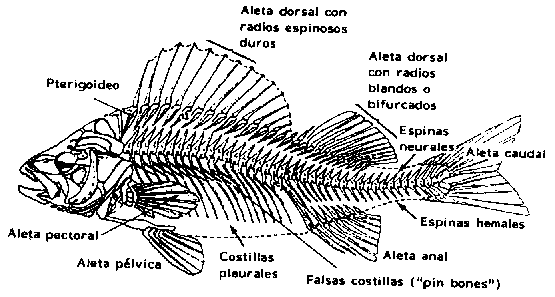
Siendo un vertebrado, el pez tiene columna vertebral y cráneo cubriendo la masa cerebral. La columna vertebral se extiende desde la cabeza hasta la aleta caudal y está compuesta por segmentos (vértebras). Estas vértebras se prolongan dorsalmente para formar las espinas neurales y en la región del tronco tienen apófisis laterales que dan origen a las costillas (Figura 2.1). Estas costillas son estructuras cartilaginosas u óseas en el tejido conectivo (miocomata) y ubicadas entre los segmentos musculares (miotomas) (véase también la Figura 2.2). Por lo general, hay también un número correspondiente de costillas falsas o "pin bones" ubicadas más o menos horizontalmente y hacia el interior del músculo. Estos huesos causan problemas importantes cuando el pescado se ha fileteado o ha sido preparado de otra manera para alimento.
Figura 2.1 Esqueleto del pez (Eriksson y Johnson, 1979)
Anatomía del músculo y su función
La anatomía del músculo del pez difiere de la anatomía de los animales terrestres, porque carece del sistema tendinoso (tejido conectivo) que conecta los paquetes musculares al esqueleto del animal. En cambio, los peces tienen células musculares que corren en paralelo, separadas perpendicularmente por tabiques de tejido conectivo (miocomata), ancladas al esqueleto y a la piel. Los segmentos musculares situados entre estos tabiques de tejido conectivo se denominan miotomas.
Figura 3.3 Musculatura esquelética del pez (Knorr, 1974)
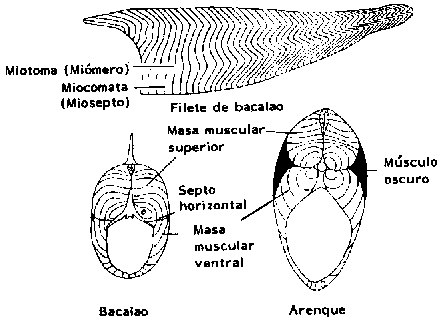
Todas las células musculares extienden su longitud total entre dos miocomatas, y corren paralelamente en el sentido longitudinal del pez. La masa muscular a cada lado del pez forma el filete. La parte superior del filete se denomina músculo dorsal y la parte inferior músculo ventral.
El largo de las células musculares del filete es heterogéneo, variando desde el final de la cabeza (anterior) hasta el final de la cola (posterior). La célula muscular más larga se encuentra en el duodécimo miotoma contado desde la cabeza y su longitud media es de alrededor 10 mm para un pescado de 60 cm de largo (Love, 1970). El diámetro de las células también varía, siendo más ancho en la parte ventral del filete.
Los miocomatas corren en forma oblicua, formando un patrón de surcos perpendiculares al eje longitudinal del pez, desde la piel hasta la espina. Esta anatomía está idealmente adaptada para permitir la flexibilidad del músculo en los movimientos necesarios para propulsar el pez a través del agua.
El tejido muscular del pez, como el de los mamíferos, está compuesto por músculo estriado. La unidad funcional, es decir, la célula muscular, consta de sarcoplasma que contiene el núcleo, granos de glucógeno, mitocondria, etc. y un número (hasta 1.000) de miofibrillas. La célula está envuelta por una cubierta de tejido conectivo denominada sarcolema. Las miofibrillas contienen proteínas contráctiles, actina y miosina. Estas proteínas o filamentos están ordenados en forma alternada muy característica, haciendo que el músculo parezca estriado en una observación microscópica (Figura 3.4).
Generalmente el tejido muscular del pez es blanco pero, dependiendo de la especie, muchos presentan cierta cantidad de tejido oscuro de color marrón o rojizo. El músculo oscuro se localiza exactamente debajo de la piel a lo largo del cuerpo del animal.
La proporción entre músculo oscuro y músculo blanco varía con la actividad del pez. En los pelágicos, es decir, especies como el arenque y la caballa, que nadan más o menos en forma continua, hasta el 48 por ciento de su peso puede estar constituido por músculo oscuro (Love, 1970). En los peces demersales, o sea, especies que se alimentan en el fondo del mar y se mueven sólo periódicamente, la cantidad de músculo oscuro es muy pequeña.
Hay muchas diferencias en la composición química de los dos tipos de músculo, siendo algunas de las más notables el alto contenido de lípidos y hemoglobina presentes en el músculo oscuro. Desde el punto de vista tecnológico, el alto contenido de lípidos del músculo oscuro resulta importante debido a los problemas asociados con la rancidez.
El color rojizo de la carne del salmón y la trucha de mar, no se origina a partir de la mioglobina sino que es debido a un carotenoide rojo, la astaxantina. La función de este pigmento no está claramente establecida, pero se ha propuesto que el carotenoide podría actuar como antioxidante. Además, su acumulación en el músculo puede funcionar como un depósito de pigmento, necesario durante el desove cuando el macho desarrolla una fuerte coloración rojiza en la piel y la hembra transporta carotenoides dentro de los huevos. El apropiado desarrollo después de la fertilización parece depender fuertemente de la cantidad de carotenoides. Se observa claramente que el color del músculo de los salmónidos se desvanece durante el desove.
El pez no sintetiza astaxantina y, por lo tanto, depende de la ingesta del pigmento a través del alimento. Algunos salmónidos viven en aguas donde la presa natural no contiene mucho carotenoide, por ejemplo el Mar Báltico, dando como resultado una coloración menos rojiza del músculo en comparación con los salmónidos de otras aguas. Esto puede ser tomado como una indicación de que la función fisiológica propuesta para la astaxantina en salmónidos, explicada en el párrafo anterior, resulte ser menos importante.
En la acuicultura del salmón, astaxantina es incluida en la alimentación, dado que el color rojo de la carne es uno de los más importantes criterios de la calidad para esta especie.
La contracción muscular comienza cuando un impulso nervioso libera Ca++ del retículo sarcoplasmático y lo lleva a las miofibrillas. Cuando la concentración de Ca++ aumenta en las enzimas activas situadas en el filamento de la miosina, la enzima ATP-asa se activa. Esta ATP-asa degrada el ATP que se encuentra entre los filamentos de actina y miosina, originando liberación de energía. La mayor parte de la energía es utilizada como energía de contracción, haciendo que los filamentos de actina se deslicen entre los filamentos de miosina, a modo de enchufe, con lo cual la fibra muscular se contrae. Cuando la reacción se invierte (o sea, cuando el Ca++ es impulsado a su lugar de origen, la actividad contráctil de la ATP-asa se detiene y permite que los filamentos se deslicen pasivamente recuperando cada uno su estado inicial), el músculo se relaja.
La fuente de energía para la generación de ATP en el músculo blanco es el glucógeno, mientras que en el músculo oscuro también puede ser obtenida a partir de los lípidos. La mayor diferencia, radica en que el músculo oscuro posee muchas más mitocondrias que el músculo blanco, permitiéndole al músculo oscuro operar extensivamente un metabolismo de energía aeróbico, resultando en la producción de CO2 y H2O como productos finales. El músculo blanco, genera la energía principalmente mediante el metabolismo anaeróbico, acumulando ácido láctico, el cual debe ser transportado al hígado para su posterior metabolización. Además, se ha reportado que el músculo oscuro posee funciones similares a las funciones encontradas en el hígado.
La diferencia entre los patrones metabólicos encontrados en los dos tipos de músculos indica que el músculo blanco está perfectamente adaptado para movimientos súbitos, fuertes y cortos; mientras que el músculo oscuro está diseñado para movimientos continuos aunque no tan fuertes.
Luego de la muerte, cesan las funciones bioquímicas y fisicoquímicas regulatorias que operan en el animal vivo y se agotan las fuentes de energía del músculo. Cuando el nivel de ATP alcanza su mínimo, los filamentos de miosina y actina quedan unidos en forma irreversible, produciéndose el rigor mortis. Este fenómeno se describe más adelante en el Capítulo 5.
El sistema cardiovascular
El sistema cardiovascular es de considerable interés para el tecnólogo pesquero dado que en algunas especies es importante desangrar el pescado (eliminar la mayor parte de la sangre) después de la captura.
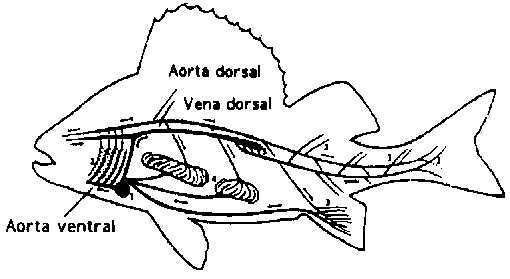
El corazón del pez está diseñado para una circulación simple (Figura 3.5). En los peces óseos el corazón consiste de dos cámaras consecutivas que bombean sangre venosa hacia las branquias, vía la aorta ventral.
Figura 3.5 Circulación de la sangre en el pez (Eriksson y Johnson, 1979)
|
Notas: |
1. El corazón bombea sangre hacia las branquias. |
|
|
2. La sangre es aireada en las branquias. |
|
|
3. La sangre arterial es dispersada dentro de los capilares, donde tiene lugar la transferencia de oxígeno y nutrientes al tejido circundante. |
|
|
4. Los nutrientes del alimento ingerido son absorbidos del intestino y transportados al hígado y posteriormente dispersados en la sangre a lo lardo de todo el cuerpo. |
|
|
5. En los riñones la sangre es "purificada" y los productos de desecho son excretados por vía urinaria. |
Después de airearse en las branquias, la sangre arterial es recogida en la aorta dorsal que corre exactamente debajo de la columna vertebral y desde aquí es dispersada en el interior de los diferentes tejidos por medio de los capilares. La sangre venosa retoma al corazón corriendo por venas de tamaño cada vez mayor (la mayor es la vena dorsal, que también se encuentra debajo de la columna vertebral). Todas las venas se juntan en un sólo vaso sanguíneo antes de entrar al corazón. El volumen total de sangre en el pez fluctúa entre el 1,5 y el 3,0 por ciento del peso del animal. La mayor parte está localizada en los órganos internos, mientras que el tejido muscular, que constituye dos tercios del peso corporal, contiene sólo el 20 por ciento del volumen de sangre. Esta distribución no cambia durante el movimiento del pez porque el músculo blanco, en particular, no está muy vascularizado.
Durante la circulación de la sangre, la presión de la misma cae desde unos 30 mg Hg en la aorta ventral hasta O cuando entra en el corazón (Randall, 1970). La presión sanguínea derivada de la actividad bombeadora del corazón, disminuye considerablemente después que la sangre ha pasado a través de las branquias. La contracción del músculo es importante en el bombeo de la sangre de regreso al corazón; el reflujo es impedido por un sistema de válvulas apareadas que se encuentran dentro de las venas.
Evidentemente, la circulación simple de la sangre en el pez es fundamentalmente diferente del sistema que presentan los mamíferos (Figura 3.6), donde la sangre pasa a través del corazón dos veces y es impulsada hacia el cuerpo a alta presión debido a las contracciones del corazón.
Figura 3.6 Circulación de la sangre en peces y mamíferos (Eriksson y Johnson, 1979)
El corazón del pez no representa un papel importante en impulsar la sangre de regreso al corazón desde los capilares. Esto ha sido confirmado en un experimento donde se analizó el efecto de diferentes procedimientos de desangrado sobre el color de filetes de bacalao. No se encontraron diferencias independientemente de la técnica de desangrado empleada: ya sea cortando delante o detrás del corazón antes de eviscerar, o sin haber efectuado ningún tipo de corte antes del sacrificio.
En algunas pesquerías, la operación de desangrado es muy importante porque se desea obtener filetes blancos uniformes. Para lograr esto, un número de países ha recomendado que el pescado se deje desangrar por un período (15-20 minutos) previo al inicio del eviscerado. Esto significa que las operaciones de desangrado y eviscerado deben efectuarse en forma separada y deben proporcionarse arreglos especiales (tanques de desangrado) en la cubierta. Esto complica el proceso de trabajo (dos operaciones en vez de una), ocasiona consumo adicional de tiempo para el pescador e incrementa la demora antes de enfriar el pescado.
Además, se requiere de espacio extra a bordo de la embarcación o de lo contrario la cubierta permanece constantemente congestionada.
Algunos investigadores han cuestionado la necesidad de manipular el pescado mediante un procedimiento de dos etapas, involucrando un período especial de desangrado (Botta et al., 1986; Huss y Asenjo, 1977a; Valdimarsson et al., 1984). Parece existir un consentimiento general en relación con lo siguiente:
· El tiempo que el pescado permanece a bordo antes de las operaciones de desangrado/eviscerado, afecta mucho más el desangrado que las propias operaciones de desangrado/eviscerado.· Se obtiene un mejor desangrado si el pescado se corta estando vivo, pero es de mayor importancia cortar el pescado antes de que entre en rigor mortis, dado que son las contracciones del músculo las que fuerzan la sangre a salir de los tejidos.
También existe desacuerdo en cuanto al método de corte. Huss y Asenjo (1977a) encontraron un mejor desangrado cuando se emplea un corte profundo en la garganta incluyendo la aorta dorsal, pero esto no fue confirmado en el trabajo de Botta et al. (1986). Este último, también recomienda incluir un período de desangrado (procedimiento de dos etapas) cuando se manipula pescado vivo (capturado con redes de encierro, trampas, redes de cerco, entre otros), mientras que Valdimarsson et al. (1984) encontraron que la calidad del bacalao muerto (4 horas después de ser llevado a bordo) mejoraba ligeramente usando el procedimiento de dos etapas. Sin embargo, es necesario señalar que el efecto del desangrado debe ser considerado en relación con las ventajas de un procedimiento de manipulación adecuado, mediante un enfriamiento rápido y eficiente de las capturas.
La decoloración de los filetes, también puede ser el resultado de una manipulación inadecuada durante la captura o mientras el pez continúa vivo. Maltrato físico en la red (prolongado tiempo de arrastre, grandes volúmenes de capturas) o en la cubierta (pescadores caminando sobre el pescado o arrojando cajas, contenedores y otros artículos sobre el pescado) puede causar contusiones, ruptura de los vasos sanguíneos y sangramiento dentro del tejido muscular (hematomas).
La aplicación de fuertes presiones sobre el pescado muerto, cuando la sangre está coagulada (por ejemplo: sobrellenando las cajas con pescado) no causa decoloración, pero el pescado puede sufrir serias pérdidas de peso.
Otros órganos
En cuanto a los otros órganos, sólo las huevas y el hígado representan un papel importante como productos comestibles. Sus tamaños dependen de la especie y varían con el ciclo biológico, la alimentación y la estación del año. En el bacalao, el peso de las huevas varía desde un pequeño porcentaje hasta el 27 por ciento del peso corporal y el peso del hígado oscila entre el 1 y el 4,5 por ciento. Además, la composición puede cambiar y el contenido de grasa del hígado puede variar entre el 15 y el 75 por ciento, el valor más alto se ha encontrado en la época de otoño (Jangaard et al., 1967).
Durante el crecimiento, aumenta el tamaño de cada célula muscular en lugar de su número. También la proporción del tejido conectivo se incrementa con la edad.
La mayor parte de los peces llegan a su madurez sexual cuando han alcanzado cierto tamaño, característico para cada especie y no está directamente correlacionado con la edad. En general este tamaño crítico se alcanza antes en los machos que en las hembras. Como la velocidad de crecimiento disminuye una vez que el pez alcanza su madurez, a menudo resulta una ventaja económica criar hembras en acuicultura.
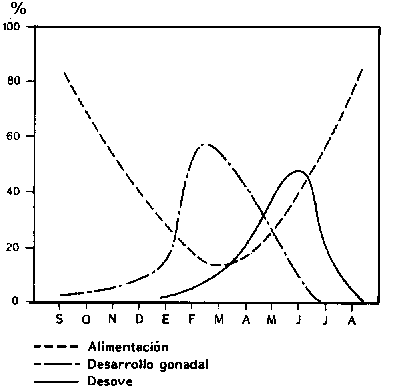
Durante todo el año, el pez sexualmente maduro gasta energía en el fortalecimiento de sus gónadas (huevas y esperma). Este desarrollo de las gónadas provoca el agotamiento de las reservas de proteínas y lípidos, porque se lleva a cabo durante un período de escasa o ninguna alimentación (Figura 3.7).
Figura 3.7 Relación entre el ciclo de alimentación (porcentaje de muestras con estómagos llenos) y el ciclo reproductivo (desarrollo de gónadas), porcentaje de pescados con madurez de gónadas (desove, porcentaje de pescado maduro) del eglefino (Melanogrammus aeglefinus). Debe notarse que el desarrollo de las gónadas ocurre mientras el pez está hambriento (Hoar, 1957).
En el bacalao del Mar del Norte se encontró que, antes del desove, el contenido de agua en el músculo aumenta (Figura 3.8) mientras que el contenido de proteínas disminuye. En casos extremos, el contenido de agua en un bacalao muy grande puede llegar a ser el 87 por ciento de su peso corporal antes del desove (Love, 1970).
Figura 3.8 Contenido de agua en el músculo de bacalao (Gadus morhua) (Love, 1970). La extensión del período de desove varía mucho entre las diferentes especies. La mayor parte de ellas tienen una marcada periodicidad estacional (Figura 3.7) mientras que algunas presentan los ovarios maduros casi todo el año.
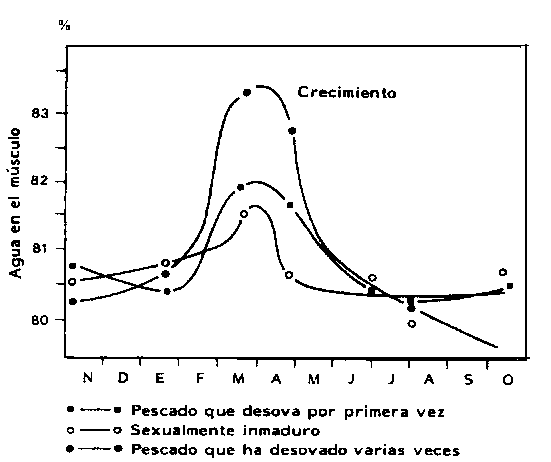
El agotamiento de las reservas del pez durante el desarrollo de las gónadas puede ser muy grave, especialmente en los casos en que la reproducción se combina con la migración hacia áreas de alimentación. Algunas especies, como por ejemplo el salmón del Pacífico (Oncorhyncus spp.), la anguila (Anguilla anguilla) y otras, migran sólo una vez, después de lo cual su estado fisiológico se deteriora en tal forma que las lleva a la muerte. Esto es debido en parte a que dichas especies no se alimentan durante la migración. Tal es el caso del salmón que puede perder durante la migración y reproducción hasta el 92 por ciento de sus lípidos, el 72 por ciento de sus proteínas y el 63 por ciento de su contenido de cenizas (Love, 1970).
Contrariamente a esto, otras especies de peces son capaces de recuperarse completamente, después de varios años de desove. El bacalao del Mar del Norte vive cerca de ocho años ante de que el desove sea causa de su muerte y otras especies pueden vivir mucho más (Cushing, 1975). En otros tiempos era usual encontrar arenques (Clupea harengus) de 25 años de edad en el Mar de Noruega, y sollas (Pleuronectes platessa) de hasta 35 años. Uno de los peces más viejos encontrados ha sido un esturión (Acipenser sturio) del Lago Winerebajo, en Wisconsin. De acuerdo con el número de anillos en los otolitos, su edad sobrepasaba los 100 años.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}