![]()
![]()
![]()
5.1 Cambios sensoriales
5.2 Cambios autolíticos
5.3 Cambios bacteriológicos
5.4 Oxidación e hidrólisis de lípidos
Los cambios sensoriales son los que percibimos a través de los sentidos, por ejemplo, apariencia, olor, textura y sabor.
Cambios en el pescado fresco crudo
Los primeros cambios sensoriales del pescado durante el almacenamiento están relacionados con la apariencia y la textura. El sabor característico de las especies normalmente se desarrolla durante los dos primeros días de almacenamiento en hielo.
El cambio más dramático es el ataque del rigor mortis. Inmediatamente después de la muerte el músculo del pescado está totalmente relajado, la textura flexible y elástica generalmente persiste durante algunas horas y posteriormente el músculo se contrae. Cuando se toma duro y rígido, todo el cuerpo se vuelve inflexible y se dice que el pescado está en rigor mortis. Esta condición generalmente se mantiene durante uno o más días y luego se resuelve el rigor. La resolución del rigor mortis hace que el músculo se relaje nuevamente y recupere la flexibilidad, pero no la elasticidad previa al rigor. La proporción entre el comienzo y la resolución del rigor varía según la especie y es afectada por la temperatura, la manipulación, el tamaño y las condiciones físicas del pescado (Cuadro 5.1).
El efecto de la temperatura sobre el rigor no es uniforme. En el caso del bacalao, las altas temperaturas ocasionan un rápido comienzo del rigor y un rigor mortis bastante fuerte. Esto debe ser evitado, dado que las fuertes tensiones producidas por el rigor pueden causar "desgajamiento", es decir, debilitamiento del tejido conectivo y posterior ruptura del filete.
Generalmente se acepta que el comienzo y la duración del rigor mortis resultan más rápido a mayor temperatura, pero se ha observado en ciertas especies tropicales el efecto opuesto de la temperatura, en relación con el comienzo del rigor. Resulta evidente que en estas especies el inicio del rigor se acelera a la temperatura de 0 °C en comparación con 10 °C, lo cual muestra buena correlación con la estimulación de los cambios bioquímicos a 0 °C (Poulter et al., 1982; Iwamoto et al,. 1987). Sin embargo, una explicación para esto ha sido sugerida por Abe y Okuma (1991), quienes han demostrado que el comienzo del rigor mortis en la carpa (Cyprinus carpió) depende de la diferencia entre la temperatura del mar y la temperatura de almacenamiento. Cuando esta diferencia es grande, el rigor se inicia a menor tiempo y viceversa.
El rigor mortis se inicia inmediatamente o poco después de la muerte, en el caso de peces hambrientos y cuyas reservas de glucógeno están agotadas, o en peces exhaustos. El método empleado para aturdir y sacrificar el pez también influye en el inicio del rigor. El aturdimiento y sacrificio por hipotermia (el pez es muerto en agua con hielo) permite obtener el más rápido inicio del rigor, mientras que un golpe en la cabeza proporciona una demora de hasta 18 horas (Azam et al., 1990; Proctor et al., 1992).
El significado tecnológico del rigor mortis es de mayor importancia cuando el pescado es fileteado antes o durante el rigor. Durante el rigor el cuerpo del pescado está completamente rígido; el rendimiento del fileteado resulta muy bajo y una manipulación tosca puede causar el desgarramiento de los filetes. Si los filetes son removidos del hueso antes del rigor, el músculo puede contraerse libremente y se encogerá al comenzar el rigor. El músculo oscuro puede encogerse hasta un 52 por ciento y el músculo blanco hasta un 15 por ciento de su longitud original (Buttkus, 1963). Si el pescado es cocido antes del rigor, la textura será muy suave y pastosa. Por el contrario, la textura es dura pero no seca cuando el pescado es cocido durante el rigor. Posterior al rigor la carne se toma firme, suculenta y elástica.
Cuadro 5.1 Comienzo y duración del rigor mortis en algunas especies de pescado
|
Especie |
Condición |
Temperatura (°C) |
Tiempo desde la muerte hasta el inicio del rigor (horas) |
Tiempo desde la muerte hasta el final del rigor (horas) |
|
Bacalao (Gadus morhua)
|
exhausto |
0 |
2-8 |
20-65 |
|
exhausto |
10-12 |
1 |
20-30 | |
|
exhausto |
30 |
0,5 |
1-2 | |
|
no exhausto |
0 |
14-15 |
72-96 | |
|
Mero (Epinephelus malabaricus) |
no exhausto |
2 |
2 |
18 |
|
Tilapia azul (Areochromis aureus) |
exhausto |
0 |
1 |
|
|
no exhausto |
0 |
6 |
| |
|
Tilapia (Tilapia mossambica) pequeña 60g |
no agotado |
0-2 |
2-9 |
26,5 |
|
Granadero (Macrourus whitson) |
exhausto |
0 |
<1 |
35-55 |
|
Anchoita (Engraulis anchoita) |
exhausto |
0 |
20-30 |
18 |
|
Solla (Pleuronectes platessa) |
exhausto |
0 |
7-11 |
54-55 |
|
Carbonero (Pollachius virens) |
exhausto |
0 |
18 |
110 |
|
Gallineta nórdica (Sebastes spp.) |
exhausto |
0 |
22 |
120 |
|
Lenguado japonés (Paralichthys olivaceus) |
|
0 |
3 |
>72 |
|
|
5 |
12 |
>72 | |
|
|
10 |
6 |
72 | |
|
|
15 |
6 |
48 | |
|
|
20 |
6 |
24 | |
|
Carpa (Cyprinus carpio) |
|
0 |
8 |
|
|
|
10 |
60 |
| |
|
|
20 |
16 |
| |
|
exhausto |
0 |
1 |
| |
|
no exhausto |
0 |
6 |
|
FUENTES: Hwang et al., 1991; Iwamoto et al., 1987; Korhonen et al., 1990; Nakayama et al., 1992; Nazir y Magar, 1963; Partmann, 1965; Pawar y Magar, 1965; Stroud, 1969; Trueco et al., 1982.
De los pescados enteros y de los filetes congelados pre-rigor, pueden obtenerse buenos productos si se descongelan cuidadosamente a baja temperatura. De esta forma, se da tiempo para que pase el rigor mortis mientras el músculo continúa congelado.
La evaluación sensorial del pescado crudo en mercados y sitios de desembarque se efectúa mediante la evaluación de la apariencia, textura y olor. Los atributos sensoriales del pescado crudo se enumeran en el Cuadro 5.2. La mayoría de los sistemas de puntuación están basados en los cambios que se producen durante el almacenamiento en hielo derretido. Debe recordarse que los cambios característicos varían dependiendo del método de almacenamiento. La apariencia del pescado almacenado en condiciones de enfriamiento sin hielo no cambia tanto en relación con el pescado en hielo, pero su deterioro es más rápido y se hace necesario efectuar una evaluación sensorial del pescado cocido. Por consiguiente, es esencial conocer la historia tiempo/temperatura del pescado al momento del desembarco.
Los cambios sensoriales característicos en el pescado post mortem varían considerablemente dependiendo de la especie y el método de almacenamiento. Una descripción general ha sido proporcionada por la Unión Europea (antes Comunidad Económica Europea) en la guía para evaluación de la calidad del pescado, como se muestra en el Cuadro 5.2. La escala sugerida está numerada de O a 3, donde 3 es la mejor calidad.
La Asociación de Tecnólogos Pesqueros de Europa Occidental (del inglés: West European Fish Technologists' Association) ha recopilado un glosario políglota de olores y sabores, que también puede ser de mucha utilidad cuando se buscan términos para describir la frescura del pescado en una evaluación sensorial (Howgate et al., 1992 (Apéndice C)).
Cambios en la calidad comestible
Cuando se requiere un criterio de calidad durante el almacenamiento del pescado refrigerado, se puede llevar a cabo una evaluación sensorial del pescado cocido. Algunos de los atributos para el pescado y los mariscos cocidos, se mencionan en el Cuadro 5.2. Se puede detectar un patrón característico del deterioro del pescado almacenado en hielo, el cual puede ser dividido en las cuatro fases siguientes:
Fase 1 El pescado es muy fresco y tiene un sabor a algas marinas, dulce y delicado. El sabor puede ser muy ligeramente metálico. En el bacalao, el eglefino, la merluza, el merlán y el lenguado, el sabor dulce se hace más pronunciado a los 2-3 días de la captura.Fase 2 Hay una pérdida del olor y del gusto característicos. La carne es neutral pero no tiene olores extraños. La textura se mantiene agradable.
Fase 3 Aparecen signos de deterioro y, dependiendo de la especie y del tipo de deterioro (aeróbico o anaeróbico), se producen una serie de compuestos volátiles de olor desagradable. Uno de estos compuestos volátiles puede ser la trimetilamina (TMA) derivada de la reducción bacteriana del oxido de trimetilamina (OTMA). La TMA tiene un olor a "pescado" muy característico. Al inicio de esta fase pueden aparecer olores y sabores ligeramente ácidos, afrutados y ligeramente amargos, especialmente en. peces grasos. En los últimos estadios de esta fase se desarrollan olores nauseabundos, dulces, como a col, amoniacales, sulfurosos y rancios. La textura se toma suave y aguada, o dura y seca.
Fase 4 El pescado puede caracterizarse como deteriorado y pútrido.
Cuadro 5.2 Clasificación de la frescura: Council Regulation (EEC) No 103/76 OJ No L20 (28 de enero de 1976) (EEC, 1976)
|
Criterio | ||||
|
Partes del pescado inspeccionadas |
Puntuación | |||
|
3 |
2 |
1 |
0 | |
|
Apariencia | ||||
|
Piel |
Pigmentación brillante e iridiscente, decoloraciones ausentes, mucus transparente y acuoso |
Pigmentación brillante pero no lustrosa |
Pigmentación en vías de descolorase y empañarse. |
Pigmentación mate¹ |
|
Ojos |
Convexos (salientes) |
Convexos y ligeramente hundidos |
Planos |
Cóncavo en el centro1 |
|
Córnea transparente |
Córnea ligeramente opalescente |
Córnea opalescente |
Córnea lechosa | |
|
Pupila negra y brillante |
Pupila negra y apagada |
Pupila opaca |
Pupila gris | |
|
Branquias |
Color brillante |
Menos coloreadas |
Descolorándose |
Amarillentas1 |
|
Mucus ausente |
Ligeros trazos de mucus |
Mucus opaco |
Mucus lechoso | |
|
Carne (corte del abdomen) |
Azulada, translúcida, uniforme, brillante |
Aterciopelada, cerosa, empañada |
Ligeramente opaca |
Opaca1 |
|
Sin cambios en el color original |
Ligeros cambios en el color |
|
| |
|
Color (a lo largo de la columna vertebral) |
No coloreada |
Ligeramente rosa |
Rosa |
Rojo1 |
|
Organos |
Riñones y residuos de otros órganos deben ser de color rojo brillante, al igual que la sangre dentro de la aorta |
Ríñones y residuos de otros órganos deben ser de color rojo empañado; la sangre comienza a decolorarse |
Ríñones, residuos de otros órganos y sangre presentan un color rojo pálido |
Ríñones, residuos de otros órganos y sangre presentan un color pardusco |
|
Condición | ||||
|
Carne |
Firme y elástica |
Menos elástica |
Ligeramente blanda (flácida), menos elástica |
Suave (flácida)1 Las escamas se desprenden fácilmente de la piel, la superficie surcada tiende a desmenuzarse |
|
Superficie uniforme |
|
Cerosa (aterciopelada) y superficie empañada |
| |
|
Columna vertebral |
Se quiebra en lugar de separarse de la carne |
Adherida |
Ligeramente adherida |
No está adherida1 |
|
Peritoneo |
Completamente adherido a la carne |
Adherido |
Ligeramente adherido |
No está adherido¹ |
|
Olor | ||||
|
Branquias, piel, cavidad abdominal |
A algas marinas |
No hay olor a algas marinas, ni olores desagradables |
Ligeramente ácido |
Acido¹ |
1O en un estado de deterioro más avanzado
Una escala numerada puede ser usada para la evaluación sensorial del pescado cocido según se muestra en la Figura 5.1. La escala está numerada del 0 al 10, donde 10 indica absoluta frescura, 8 buena calidad y 6 un pescado con sabor neutro (insípido). El nivel de rechazo es 4. Usando la escala según la puntuación señalada, el gráfico adquiere forma de "S" indicando una rápida degradación del pescado durante la primera fase, menor tasa en las fases 2 y 3, y finalmente una alta variación cuando el pescado se descompone.
Figura 5.1 Cambios en la calidad comestible del bacalao en hielo (0°C) (Huss 1976)
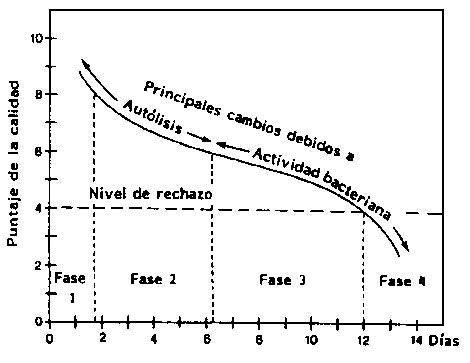
Otras escalas también pueden ser empleadas y cambiar la forma del gráfico. Sin embargo, es importante entender la clase de resultados deseados en el análisis sensorial, a fin de efectuar las preguntas adecuadas a los evaluadores sensoriales.
Autólisis significa "auto-digestión". Se sabe desde hace muchos años que existen por lo menos dos tipos de deterioro en el pescado: bacteriano y enzimático. Uchyama y Ehira (1974), demostraron que en el bacalao y en el atún aleta amarilla, los cambios enzimáticos relativos a la frescura del pescado precedían y no guardaban relación con los cambios de la calidad microbiológica. En algunas especies (calamar, arenque), los cambios enzimáticos preceden y por lo tanto predominan al deterioro del pescado refrigerado. En otros la autólisis, sumada al proceso microbiano, contribuye en diferentes grados a la pérdida general de la calidad.
Producción de energía en el músculo post mortem
Al momento de la muerte, el suministro de oxígeno al tejido muscular se interrumpe porque la sangre deja de ser bombeada por el corazón y no circula a través de las branquias donde, en los peces vivos, es enriquecida con oxígeno. Dado que el oxígeno no está disponible para la respiración normal, se restringe la producción de energía a partir de los nutrientes ingeridos. La Figura 5.2 ilustra la ruta normal para la producción de energía muscular en la mayoría de los peces teleósteos vivos (peces óseos con aletas). El glucógeno (carbohidrato de almacenamiento) o las grasas son oxidadas o "quemadas" por las enzimas del tejido, en una serie de reacciones las cuales finalmente producen dióxido de carbono (CO2), agua y adenosina trifosfato (ATP), un compuesto orgánico rico en energía. Este tipo de respiración se efectúa en dos etapas: una anaeróbica y otra aeróbica. La última depende de la continua presencia del oxígeno (O2), sólo disponible en el sistema circulatorio. La mayoría de los crustáceos son capaces de respirar fuera del ambiente acuático por períodos limitados de tiempo, mediante absorción del oxígeno atmosférico.
Figura 5.2 Descomposición aeróbica y anaeróbica del glucógeno en el músculo del pescado
La Figura 5.2 también ilustra el hecho de que en condiciones de anaerobiosis, el ATP puede ser sintetizado a través de otras dos importantes rutas a partir de la creatina fosfato o la arginina fosfato. La primera fuente de energía está restringida al músculo de los vertebrados (peces teleósteos), mientras que la segunda es característica de algunos invertebrados como los cefalópodos (calamar y pulpo). En cualquiera de los casos, la producción de ATP cesa en cuanto se agotan la creatina fosfato o la arginina fosfato. Resulta interesante notar que la octopina es el producto final del metabolismo anaeróbico de los cefalópodos y no es de naturaleza ácida (a diferencia del lactato), así que cualquier cambio en el pH post mortem, en este tipo de animales, no está relacionado con la producción de ácido láctico a partir del glucógeno.
Para la mayoría de los peces teleósteos, la glucólisis es la única ruta posible para la producción de energía en cuanto el corazón deja de latir. Este proceso, más ineficiente, genera principalmente ácido láctico y ácido pirúvico como productos finales. Además, mediante la glucólisis se producen dos moles de ATP por cada mol de glucosa, en comparación con los 36 moles de ATP producidos por cada mol de glucosa si los productos glucolíticos finales son oxidados aeróbicamente en la mitocondria del animal vivo. Así, después de la muerte, el músculo anaeróbico no puede mantener su nivel normal de ATP, y cuando el nivel intracelular declina de 7-10 m moles/g a £ 1,0 m moles/g de tejido, el músculo entra en rigor mortis. La glucólisis post mortem resulta en la acumulación de ácido láctico, con la concomitante disminución del pH en el músculo. En el bacalao, el pH disminuye desde 6.8 hasta un pH extremo de 6.1-6.5. En algunas especies de pescado, el pH final puede ser menor: en caballas grandes, el pH extremo en el rigor puede llegar a ser tan bajo como 5.8-6.0, y en atunes e hipoglosos se han encontrado valores tan bajos como 5.4-5.6. Sin embargo, estos niveles tan bajos de pH no son frecuentes en teleósteos marinos. Estos pH rara vez son tan bajos como los observados en el músculo post mortem de mamíferos. Por ejemplo, el pH del músculo de vacuno generalmente disminuye a niveles de 5.1 durante el rigor mortis. La cantidad de ácido láctico producido está relacionada con la cantidad de carbohidrato almacenado (glucógeno) en el tejido vivo. En general, el músculo de pescado contiene un nivel relativamente bajo de glucógeno, comparado con los mamíferos y por esta razón se genera mucho menos ácido láctico después de la muerte. También el estado nutricional del pez, la cantidad y grado de agotamiento al momento de la muerte, tienen un efecto dramático en los niveles de glucógeno almacenado y consecuentemente en el pH post mortem final. Como regla, el pescado bien descansado y bien alimentado contiene más glucógeno que el pescado exhausto y hambriento. En un estudio reciente de la locha japonesa (Chiba et al., 1991), se demostró que sólo minutos de agotamiento antes de la captura, ocasionaban una disminución de 0.50 unidades de pH en 3 horas, en comparación con peces no sometidos a agotamiento, en los cuales el pH disminuyó en sólo 0,10 unidades durante el mismo período de tiempo. Además, los mismos autores demostraron que el desangrado del pescado disminuye significativamente la producción de ácido láctico post mortem.
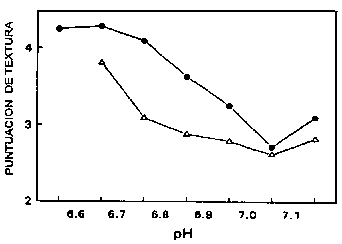
La disminución post mortem en el pH del músculo de pescado tiene un efecto en las propiedades físicas del músculo. A medida que el pH disminuye, se reduce la carga neta de la superficie de las proteínas musculares, causando su desnaturalización parcial y disminuyendo su capacidad de enlazar agua. El músculo en estado de rigor mortis pierde su humedad cuando es cocido y resulta particularmente inadecuado para un procesamiento posterior que involucre calentamiento, puesto que la desnaturalización por calor incrementa la pérdida de agua. La pérdida de agua tiene un efecto perjudicial en la textura del músculo; ha sido demostrado por Love (1975) que existe una relación inversamente proporcional entre la dureza del músculo y el pH, donde los niveles inaceptables de dureza (y pérdidas de agua por cocción) ocurren a menores niveles de pH (Figura 5.3).
Figura 5.3. Relación entre la textura del músculo de bacalao y el pH, adaptado de Love (1975). Los puntos negros se refieren a pescado capturado en St. Kilda, Océano Atlántico, mientras que los triángulos se refieren a pescado capturado en Fyllas Bank, Estrecho de Davis.
Autólisis y catabolismo de nucleótidos
Como se mencionó anteriormente, el rigor mortis se establece cuando el nivel de ATP en el músculo cae a £ 1.0 m moles/g. El ATP no es sólo una fuente de alta energía necesaria para la contracción muscular de los animales vivos, sino que también proporciona plasticidad al músculo. La contracción muscular per se está controlada por el calcio y la enzima ATP-asa que se encuentra en cada célula muscular. Cuando los niveles de Ca+² intracelular son >1m M, la ATP-asa activada por Ca+2 reduce los niveles de ATP libre en el músculo, ocasionando la interacción entre la actina y la miosina, las principales proteínas contráctiles. Esta interacción trae como resultado la reducción del músculo, ocasionando su endurecimiento y pérdida de la flexibilidad. Durante el rigor mortis, el pescado no puede ser fileteado o procesado normalmente, porque el cuerpo está demasiado rígido para ser manipulado y generalmente retorcido, impidiendo su manipulación mediante maquinaria (véase también la Sección 3.2 sobre desangrado y Sección 5.1 sobre cambios sensoriales).
La resolución del rigor es un proceso no del todo comprendido, pero siempre ocasiona el reblandecimiento (relajación) posterior del tejido muscular y se cree está relacionado con la activación de una o más enzimas musculares presentes en el pescado, las cuales digieren ciertos componentes del complejo rigor mortis. El reblandecimiento del músculo durante la resolución del rigor (y eventualmente el proceso de deterioro) coincide con los cambios autolíticos. De estos cambios, el primero en ser reconocido de forma más o menos predecible después de la muerte fue la degradación de los compuestos relacionados con el ATP. La Figura 5.4 ilustra la degradación del ATP para formar adenosina difosfato (ADP), adenosina monofosfato (AMP), inosina monofosfato (IMP), inosina (Ino) e Hipoxantina (Hx). La degradación de los catabolitos del ATP procede de la misma forma en la mayoría de los pescados, pero la velocidad de cada reacción (de un catabolito a otro), varía enormemente entre una especie y otra, coincidentemente, progresando generalmente con el nivel percibido de deterioro según determinaciones efectuadas mediante un panel de analistas entrenados. Saito et al. (1959), fueron los primeros en observar este patrón y desarrollaron una fórmula para la frescura del pescado basada en estos cambios autolíticos:
Donde [ATP], [ADP], [AMP], [IMP], [Ino] e [Hx], representan las concentraciones relativas de estos compuestos en el músculo de pescado, medidas en diferentes períodos de tiempo durante el almacenamiento refrigerado.
El índice de frescura K proporciona una puntuación de frescura relativa, basada principalmente en los cambios autolíticos que tienen lugar durante el almacenamiento post mortem del músculo. De este modo, cuanto más alto el valor de K, menor el nivel de frescura. Desdichadamente, algunas especies de pescado, como el bacalao del Atlántico, alcanzan un valor K máximo mucho antes que la vida en anaquel, según lo determinado por jueces entrenados. Por lo tanto, K no puede ser considerado como un índice confiable de frescura para todos los peces marinos con aletas. Asimismo, la degradación de nucleótidos es sólo coincidencial con los cambios percibidos en la frescura y no está necesariamente relacionada con su deterioro, considerándose que sólo la hipoxantina (Hx) tiene un efecto directo en el sabor amargo percibido en el pescado deteriorado (Hughes y Jones, 1966). Actualmente, es ampliamente aceptado que la IMP es responsable del deseable sabor a pescado fresco, sólo presente en los productos pesqueros de alta calidad. Ninguno de los nucleótidos se considera relacionado a los cambios percibidos en la textura durante el proceso autolítico, a excepción del ATP, por supuesto, cuya disminución está asociada con el rigor mortis.
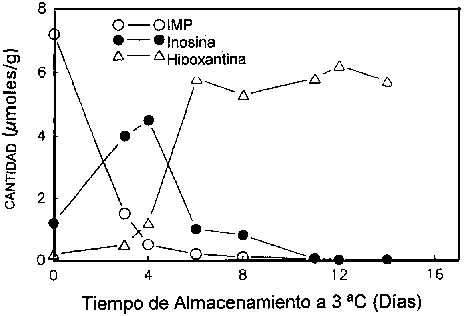
Surette et al. (1988) siguieron la autólisis de bacalao estéril y no estéril mediante los catabolitos de ATP. La velocidad de formación y descomposición del IMP fue la misma tanto en las muestras de tejido del bacalao estéril como en las del bacalao no estéril (Figuras 5.5a y 5.5b), lo cual indica que la ruta catabólica para la degradación de ATP hasta inosina es debida en su totalidad a enzimas autolíticas.
La conversión de inosina a hipoxantina se aceleró 2 días en las muestras no estériles. Esto sugiere que la nucleosida fosforilasa bacteriana (enzima 5a en la Figura 5.4) desempeña un papel principal en la producción post mortem de hipoxantina en bacalao refrigerado (véase también sección 5.3). Es interesante notar que Surette et al. (1988) no lograron recuperar nucleosida fosforilasa a partir de bacalao recién muerto, pero Surette et al. (1990) continuaron posteriormente con el aislamiento y purificación de esta enzima a partir de la bacteria Proteus, recuperada en filetes de bacalao deteriorado. Como se mencionó anteriormente, es de esperarse grandes variaciones en los patrones de la degradación de nucleótidos entre una especie y otra. Las variaciones de hipoxantina entre los diferentes tipos de pescado se muestran en la Figura 5.6. Está claro por lo tanto, que la determinación de hipoxantina no resulta de utilidad en especies como el pez espada y la gallineta nórdica.
Figura 5.5a Cambios en IMP, Ino y Hx en filetes estériles de bacalao a 3°C, adaptado de Gill (1990)
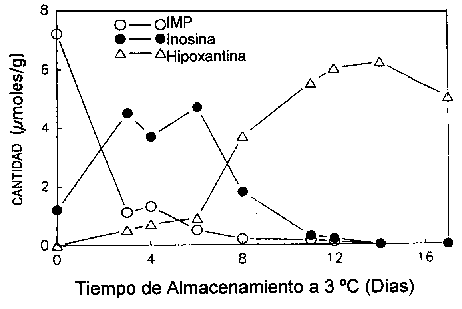
Figura 5.5b Cambios en IMP, Ino y Hx en filetes no estériles de bacalao a 3°C, adaptado de Gill (1990)
Existe poca duda en que la manipulación física acelera los cambios autolíticos en pescado refrigerado. Surette et al. (1988) reportaron que la tasa de descomposición de los nucleótidos era mayor en filetes estériles que en bacalao entero eviscerado no estéril. Esto quizá no sea sorprendente, pues se ha demostrado que muchas de las enzimas autolíticas se encuentran en discretos paquetes limitados por membranas, los cuales se rompen cuando están sujetos a abuso físico, originando la mezcla entre enzimas y sustratos. Aplastar el pescado contra el hielo o contra otros pescados puede afectar seriamente la comestibilidad y el rendimiento en el fileteado, incluso para pescados con cargas bacterianas relativamente bajas, lo cual demuestra la importancia de los procesos autolíticos. A fin de minimizar la autólisis, el pescado en hielo nunca debe ser almacenado en cajas cuya profundidad exceda los 30 cm y de igual forma es importante asegurar que las cajas no vayan apretadas unas encima de la otras. Deben ser diseñados sistemas para transportar y descargar el pescado de los barcos, que permitan evitar daño físico a los delicados tejidos.
Se han desarrollado algunos métodos rápidos para la determinación de nucleótidos individualmente o en combinaciones, incluyendo el índice de frescura. Pueden ser consultadas dos revisiones recientes (Gill, 1990, 1992).
Cambios autolíticos que involucran enzimas proteolíticas
Muchas proteasas han sido aisladas del músculo de pescado y el efecto de la descomposición proteolítica está generalmente relacionado con un extenso ablandamiento del tejido. Quizá uno de los más notables ejemplos de la proteólisis autolítica es la incidencia de vientre desgarrado (estallido de vientre) en especies pelágicas (pescado graso) como el arenque y el capelán. Este tipo de ablandamiento del tejido es más predominante durante los meses de verano, cuando los pelágicos se alimentan abundantemente, particularmente de un alimento constituido por copepodos y eufausiidos ("red feed"). Los péptidos de bajo peso molecular y los aminoácidos libres producidos por la autólisis de las proteínas no sólo disminuyen la aceptación comercial de los pelágicos. También se ha demostrado, en capelán almacenado, que la autólisis acelera el crecimiento de las bacterias del deterioro, proporcionando un medio de crecimiento superior para este tipo de organismos (Aksnes y Brekken, 1988). La inducción del deterioro bacteriano en el capelán -por autólisis- también ocasiona la descarboxilación de aminoácidos, produciendo aminas biógenas y disminuyendo significativamente el valor nutritivo del pescado. Esto es de particular importancia, puesto que la autólisis y el crecimiento bacteriano disminuyen enormemente el valor comercial de los pelágicos empleados en la fabricación de harina de pescado.
También se han encontrado carboxipeptidasas A y B, quimotripsina y tripsina, en arenque almacenado a granel para la fabricación de harina de pescado. Estudios preliminares han demostrado que la proteólisis puede ser inhibida mediante la adición de extracto de papa, el cual no sólo retarda la proteólisis sino que también disminuye el crecimiento microbiano y preserva los valores nutricionales de la harina (Aksnes, 1989).
Más recientemente, Botta et al. (1992) encontraron que la autólisis de la cavidad visceral (estallido de vientre) en el arenque estaba más relacionada con la manipulación física que con factores biológicos como el tamaño del pescado, cantidad de alimento ("red feed") en las vísceras o presencia de huevas. Particularmente se demostró que en arenque congelado/descongelado, el tiempo de descongelado a 15 °C y el tiempo del almacenamiento en hielo, tienen una influencia mucho mayor en el estallido de vientre que los factores biológicos.
Catepsinas
Si bien han sido aisladas varias enzimas proteolíticas en el tejido del pescado, han sido las catepsinas las que quizás se han descrito con mayor frecuencia. Las catepsinas son proteasas "ácidas" que usualmente se encuentran empacadas en diminutos organelos submicroscópicos llamados lisosomas. En el tejido vivo, las proteasas lisosomales se cree son responsables de la degradación proteica en las áreas de daño. De esta forma, las catepsinas están generalmente inactivas dentro del tejido vivo pero son liberadas dentro de los fluidos celulares luego de abuso físico o congelación y descongelación post mortem del músculo.
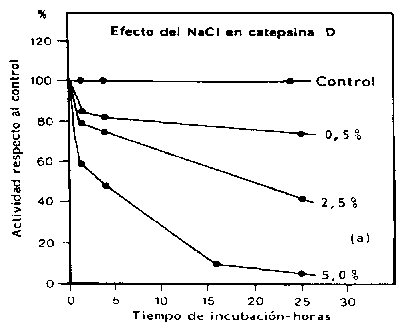
Se cree que las catepsinas D y L desempeñan un papel primordial en la degradación autolítica del tejido del pescado, dado que la mayor parte de las otras catepsinas presentan actividad en un rango relativamente estrecho de pH, demasiado bajo para tener significado fisiológico. Reddi et al. (1972) demostraron que una enzima del lenguado de invierno, se cree la catepsina D, es activa dentro de un rango de pH de 3-8 con un máximo cerca del pH 4.0, aunque no se efectuó ningún intento para confirmar la identidad de la enzima empleando un sustrato sintético o inhibidores específicos. Sin embargo, la enzima es mucho menos activa en presencia de ATP, lo cual sugiere que esta enzima estaría activa sólo en el músculo de pescado post mortem. Además, la actividad de la enzima es fuertemente inhibida en presencia de sal (Figura 5.7). Virtualmente, no hay actividad remanente después de 25 horas de incubación en una solución al 5 por ciento de cloruro de sodio. Por consiguiente, resulta improbable que la enzima de Reddi permanezca activa en productos salados.
Catepsina L ha sido implicada en el ablandamiento del músculo de salmón durante la migración por desove. Al parecer, esta enzima contribuye a la autólisis del músculo de pescado más que la catepsina D, dado que es mucho más activa a pH neutro, y se ha demostrado que digiere tanto proteínas miofibrilares (actiomiosina) como tejido conectivo. Yamashita y Konogaya (1990), obtuvieron fuerte evidencia implicando la catepsina L, antes que otras catepsinas, en el ablandamiento del salmón durante el desove. Ellos demostraron que la electroforesis de miofibrillas purificadas tratadas con catepsinas L mostraban patrones casi idénticos a los patrones de proteínas recuperadas del músculo del pescado en desove. Más aún, la actividad autolítica de la catepsina L se correlaciona muy bien con la textura del músculo, según mediciones instrumentales. La correlación linear entre la actividad de la catepsina L y la fuerza de ruptura del músculo fue excelente; r == 0.86 para tejido fresco y r = -0.95 para tejido congelado/descongelado. Es interesante notar que en todos los casos la habilidad autolítica, medida como actividad de catepsina L, resultó mayor en el tejido congelado/descongelado que en el tejido fresco. La congelación y descongelación, generalmente causan interrupciones en la membrana celular, permitiendo que las enzimas autolíticas reaccionen con su sustrato natural. La enzima y su inhibidor natural fueron estudiados posteriormente por los mismos autores (Yamashita y Konogaya, 1992). La catepsina L también ha sido asociada con la producción de un ablandamiento gelatinoso en lenguado (Toyohara et al., 1993a) y el ablandamiento incontrolable en el músculo de la merluza del Pacífico, parasitada por Myxosporidia (Toyohara et al., 1993b).
Figura 5.7 Efecto del NaCl en la actividad de la catepsina. Adaptado de Reddi et al. (1972)
Los tejidos del pescado infectado tienen poco valor comercial, pero actualmente se desconoce si es el parásito o el huésped quien secreta las enzimas proteolíticas que autolizan el músculo.
Además de su perjudicial efecto en la textura, las enzimas catepsinas inducen cambios autolíticos intencionales en productos pesqueros fermentados. Por ejemplo, se cree que las catepsinas son responsables de los principales cambios en la textura durante la fermentación del calamar japonés y la carpa Cruciana preservados en sal (Makinodan et al., 1991, 1993).
Calpainas
Un segundo grupo de proteasas intracelulares denominadas "calpainas" o "Factor Activado por Calcio" (FAC o del ingles CAF = Calcium Activated Factor) han sido recientemente asociadas con la autólisis del músculo de pescado y se les encuentra en carnes, pescados de aleta y crustáceos. La suavidad y jugosidad son probablemente las características de calidad más importantes en la carne roja. Desde hace casi un siglo se conoce que la maduración post mortem de la carne roja ocasiona el proceso de ablandamiento. Las calpainas han sido encontradas como las principales responsables de la autólisis post mortem de la carne, debido a la digestión de las proteínas de la Línea Z de las miofibrillas. Si bien el endurecimiento es rara vez un problema en el músculo no congelado, el ablandamiento debido a la autólisis es un problema serio que limita su valor comercial. Las calpainas son endopeptidasas intracelulares, cisteína y calcio dependientes; m -calpaina requiere 5-50 m M Ca+2, m-calpaina requiere 150-1000 m M Ca+2. La mayoría de las calpainas son activas a pH fisiológico, lo cual hace razonable sospechar su importancia en el ablandamiento del pescado durante el almacenamiento refrigerado.
Estudios han demostrado que en el músculo de crustáceos, las calpainas están asociadas con los cambios textuales de licuefacción inducida al músculo y digestión inespecífica generalizada de las proteínas miofibrilares. Sin embargo, las calpainas del músculo de vertebrados han demostrado ser muy específicas, degradando principalmente tropinina-T, desmina, titina y nebulina, atacando tanto actina como miosina en vertebrados (Koohmaraie, 1992). Por el contrario, las calpainas de pescado degradan miosina (específicamente la cadena pesada de la miosina) para formar un fragmento inicial con un peso molecular de aproximadamente 150 000 Da (Muramoto et al., 1989). Los mismos autores demostraron que las calpainas de pescado son mucho más activas a bajas temperaturas que las calpainas de mamíferos y las tasas de escisión son específicas de la especie, siendo más activas contra las miosinas menos estables al calor. De este modo, las especies de peces adaptadas a bajas temperaturas ambientales son más susceptibles a la autólisis por calpainas, que las especies de aguas tropicales. Aunque la calpaina ha sido identificada en distintas especies de peces incluyendo la carpa (Toyohara et al., 1985), tilapia y camarón (Wang et al., 1993), como también en atún, roncador, besugo rojo y trucha (Muramoto et al., 1989) por nombrar algunos, pocos trabajos hasta ahora demuestran una relación de "causa y efecto" entre la actividad de la calpaina y la medición instrumental de la textura.
Colagenasas
Hasta este punto, todos los cambios autolíticos post mortem descritos involucran cambios dentro de la célula muscular per se. Sin embargo, la carne de los peces teleósteos está dividida en bloques de células musculares separadas en "escamas", o miotomas, mediante tejido conectivo denominado miocomata (Figura 3.3). Cada célula muscular o fibra está rodeada por tejido conectivo que se une a la miocomata al final de la célula mediante finas fibrillas de colágeno. Durante el almacenamiento refrigerado, estas fibrillas se deterioran (Bremner y Hallett, 1985). Más recientemente, se demostró que la medición instrumental de la textura del músculo de trucha refrigerada decae a medida que se solubilizan los niveles de colágeno tipo V, presumiblemente debido a la acción de las enzimas colagenasas autolíticas (Sato et al., 1991). Son estas enzimas las que presumiblemente causan "desgajamiento", o ruptura de los miotomas, durante el almacenamiento prolongado en hielo o durante el almacenamiento por cortos períodos de tiempo, pero a elevadas temperaturas. En el bacalao del Atlántico se ha demostrado que al alcanzar los 17 °C, el desgajamiento es inevitable, debido presumiblemente a la degradación del tejido conectivo y el rápido acortamiento del músculo por la elevada temperatura durante el rigor.
También se ha demostrado que la relativa corta duración en almacén de los camarones pequeños (prawn = quisquillas), debido al ablandamiento del tejido, es ocasionada por la presencia de enzimas colagenasas (Nip et al., 1985). Se piensa que la fuente de colagenasas en las quisquillas es el hepatopáncreas (órgano digestivo).
Cambios autolíticos durante el almacenamiento en congelación
La reducción del óxido de trimetilamina (OTMA), un compuesto osmorregulatorio presente en muchos peces teleósteos marinos, se debe usualmente a la acción bacteriana (sección 5.3) pero algunas especies presentan en el tejido muscular una enzima capaz de descomponer el OTMA en dimetilamina (DMA) y formaldehído (FA):
(CH3)3NO ® (CH3)2NH + HCHO
Es importante notar que la cantidad de formaldehído producido es equivalente a la dimetilamina formada, pero su significado comercial es de mayor importancia. El formaldehído induce el entrecruzamiento de las proteínas musculares ocasionando endurecimiento del músculo y pérdida de su capacidad para enlazar agua. La enzima responsable del endurecimiento inducido por el formaldehído es la OTMA-asa, o OTMA dimetilasa y se encuentra más comúnmente en los peces gádidos (familia de los bacalaos). La mayoría de las enzimas OTMA dimetilasas reportadas hasta ahora están unidas a la membrana y se toman más activas cuando el tejido de la membrana es roto por la congelación o artificialmente, por la solubilizaron en detergentes. El músculo oscuro (rojo) presenta una mayor tasa de actividad que el músculo blanco, mientras otros tejidos como el riñón, el bazo y la vesícula biliar son extremadamente ricos de la enzima. En tal sentido, es importante que el pescado deshuesado esté completamente libre de tejidos de órganos, como el riñón de gádidos, si desea evitarse el endurecimiento durante el almacenamiento en congelación. Generalmente resulta difícil asegurar que los riñones han sido removidos antes del deshuesado mecánico, dado que este órgano en particular se localiza a lo largo de la espina y está adherido a ella. La enzima OTMA-asa ha sido aislada de la fracción microsomal en músculo de merluza (Parkin y Hultin, 1986) y de la membrana lisosomal en tejido de riñón (Gill et al; 1992). Se ha demostrado que el endurecimiento del músculo de merluza congelada se correlaciona con la cantidad de formaldehído producido, y que la tasa de producción de formaldehído es mayor a altas temperaturas de almacenamiento en congelación (Gill et al., 1979). Además, se ha demostrado que la cantidad de endurecimiento inducido por el formaldehído se intensifica por el abuso físico durante la captura, antes de la congelación, y por las fluctuaciones de la temperatura durante el almacenamiento en congelación. El medio más práctico para prevenir la producción autolítica de formaldehído es almacenando el pescado a temperaturas <-30°C, a fin de minimizar las fluctuaciones de temperatura en el almacenamiento, y evitando la manipulación tosca o la aplicación de presión física sobre el pescado antes del congelamiento. Los cambios autolíticos que afectan la comestibilidad del pescado fresco y congelado se resumen en el Cuadro 5.3. Generalmente, el factor de mayor influencia en la autólisis es la desorganización física de las células musculares. En el presente documento no se tratan las proteasas alcalinas asociadas con el ablandamiento de los productos cocidos basados en surimi. En un artículo de Kinoshita et al. (1990) se tratan Las proteasas alcalinas activadas por calor, asociadas con el ablandamiento en productos basados en surimi.
Cuadro 5.3 Resumen de los Cambios Autolíticos en el Pescado Enfriado
|
Enzima (s) |
Sustrato |
Cambios encontrados |
Prevención/Inhibición |
|
Enzimas glucolíticas |
glucógeno |
· producción de ácido láctico, disminución del
pH de los tejidos, pérdida de la capacidad de enlazar agua en el
músculo |
· el pescado debe pasar por la etapa de
rigor a temperaturas lo más cercanas a 0 °C |
|
Enzimas autolíticas, involucradas en la degradación de nucleótidos |
ATP |
· pérdida del sabor a pescado fresco, producción gradual del sabor amargo con Hx (estados finales) |
· igual que el anterior |
|
Catepsinas |
proteínas, péptidos |
· ablandamiento del tejido dificultando o impidiendo su procesamiento |
· la manipulación inadecuada el almacenamiento y la descarga |
|
Quimotripsina, tripsina carboxipeptidasas |
proteínas, péptidos |
· autólisis de la cavidad visceral en pelágicos (estallido de vientre) |
· el problema se agrava por congelación/descongelación y el almacenamiento en frío prolongado |
|
Calpaína |
proteínas miofibrilares |
· ablandamiento, ablandamiento inducido por muda en crustáceos |
· ¿remover del calcio para prevenir la activación? |
|
Colagenasas |
tejido conectivo |
· "desgajamiento" de filetes |
· la degradación del tejido conectivo está relacionada con el tiempo y temperatura de almacenamiento en refrigeración |
|
OTMA desmetilasa |
OTMA |
· endurecimiento inducido por formaldehído (gádidos almacenados en congelación) |
· temperatura de almacenamiento del pescado
< -30 °C |
La flora bacteriana en peces vivos
Los microorganismos se encuentran en todas las superficies externas (piel y branquias) y en los intestinos de los peces vivos y recién capturados. El número total de microorganismos varía enormemente, Liston (1980) establece como rango normal 102 - 107 ufc (unidades formadoras de colonias)/cm2 en la superficie de la piel. Las branquias e intestinos contienen entre 103 y 109 ufc/g (Shewan, 1962).
La flora bacteriana en pescados recién capturados depende más del medio ambiente de captura, que de la especie (Shewan, 1977). Los pescados capturados en aguas muy frías y limpias contienen menor número de microorganismos, mientras que el pescado capturado en aguas cálidas presenta recuentos ligeramente superiores. Números muy elevados, por ejemplo 107 ufc/cm2, se encuentran en pescados capturados en aguas muy contaminadas. Muchas especies diferentes de bacterias pueden ser encontradas en la superficie de los peces. Las bacterias en peces de aguas templadas son clasificadas en psicrotrófas y psicrófilas, de acuerdo al rango de su temperatura de crecimiento. Las psicrotrófas (tolerantes al frío) son bacterias capaces de crecer a 0 °C pero su óptimo es alrededor de los 25 °C. Las psicrófilas (amantes del frío) son bacterias con una temperatura máxima de crecimiento alrededor de los 20 °C y su óptimo a 15 °C (Morita, 1975). En las aguas cálidas pueden aislarse un mayor número de mesófilos. La microflora en peces de aguas templadas está dominada por bacterias psicrófilas Gram negativas con forma de bastones, pertenecientes a los géneros Pseudomonas, Moraxella, Acinetobacter, Shewanella y Flavobacterium. Miembros de las Vibrionáceas (Vibrio y Photobacterium) y Aeromonadáceas (Aeromonas spp.) son también bacterias acuáticas comunes y típicas de la flora bacteriana en pescado (Cuadro 5.4). Organismos Gram positivos como Bacillus, Micrococcus, Clostridium, Lactobacillus y coryneformes también pueden ser encontrados en distintas proporciones. Pero en general, las bacterias Gram-negativas dominan la microflora. Shewan (1977) concluyó que las bacterias Gram-positivas Bacillus y Micrococcus dominaban la microflora en pescados de aguas tropicales. Sin embargo, esta conclusión fue confrontada posteriormente por varios estudios en los cuales se encontró que la flora, en especies de peces tropicales, es muy similar a la flora en especies templadas (Acuff et al., 1984; Gram et al., 1990; Lima dos Santos 1978; Surendran et al., 1989). En algunos estudios realizados en la India se ha encontrado una microflora compuesta por Pseudomonas, Acinetobacter, Moraxella y Vibrio en pescado recién capturado (Surendran et al., 1989). Algunos autores, como Liston (1980), concluyen que la microflora de los peces tropicales a menudo contiene una carga ligeramente mayor de bacterias Gram-positivas y bacterias entéricas, pero por lo demás es similar a la flora de los peces de aguas templadas.
Las Aeromonas spp. son típicas de los peces de agua dulce, mientras que otras bacterias requieren sodio para su crecimiento y, por lo tanto, son típicas de aguas marinas. Este grupo incluye Vibrio, Photobacterium y Shewanella. Sin embargo, a pesar de que Shewanella putrefaciens se caracteriza como dependiente de sodio, también pueden aislarse cepas de S. putrefaciens, a partir de ambientes de agua dulce (DiChristina y DeLong, 1993; Gram et al; 1990; Spanggaard et al., 1993). A pesar de que S. putrefaciens ha sido aislada de aguas dulces tropicales, no resulta de importancia en el deterioro del pescado de agua dulce (Lima dos Santos, 1978; Gram, 1990).
Cuadro 5.4 Flora bacteriana de pescado capturado en aguas limpias no contaminadas
|
Gram-negativas |
Gram-positivas |
Comentarios |
|
Pseudomonas |
Bacillus |
|
|
Moraxella |
Clostridium |
|
|
Acinetobacter |
Micrococcus |
|
|
Shewanella putrefaciens |
Lactobacillus |
|
|
Flavobacterium |
Coryneformes |
|
|
Cytophaga |
|
|
|
Vibrio |
|
Vibrio y Photobacterium son típicas de aguas marinas; |
|
Photobacterium |
|
|
|
Aeromonas |
|
Aeromonas es típica de agua dulce |
En aguas contaminadas, puede encontrarse un elevado número de Enterobacteriáceas. En aguas limpias y templadas, estos organismos desaparecen rápidamente, pero se ha demostrado que Escherichia coli y Salmonella pueden sobrevivir por períodos bastante prolongados de tiempo en aguas tropicales y una vez introducidos en el ambiente, se convierten casi que en autóctonos (Fujioka et al., 1988).
La taxonomía de S. putrefaciens ha sido algo confusa. El organismo fue originalmente asociado con el grupo Achromobacter, pero posteriormente colocado en el grupo IV de Shewan Pseudomonas. Tomando como base el porcentaje de guanina + citosina (GC%) se le transfirió al género Alteromonas, pero sobre la base de la homología del 5SRNA se le clasificó en un nuevo género, Shewanella (MacDonnell y Colwell, 1985). Recientemente se ha sugerido que el género Aeromonas spp., un miembro de la familia de las Vibrionáceas, sea transferido a su propia familia, la Aeromonadáceas (Colwell et al., 1986).
Estudios japoneses, han mostrado la presencia de un número muy elevado de microorganismos en el tracto gastrointestinal del pescado, inclusive superior al de las aguas circundantes; esto indica la presencia de un nicho ecológico favorable para los microorganismos. Igualmente, Larsen et al. (1978) reportan hasta 107 ufc/g de organismos parecidos al Vibrio en el tracto intestinal del bacalao, Westerdahl et al. (1991) también aislaron un elevado número de microorganismos parecidos al Vibrio de los intestinos de lenguados. Photobacterium phosphoreum puede ser aislado de la superficie externa y también puede ser aislado en un elevado número del tracto intestinal de algunas especies de pescado (Dalgaard, 1993). Por el contrario, algunos autores consideran que la microflora del tracto gastrointestinal es meramente un reflejo del medio ambiente y de la ingesta.
Invasión microbiana
El músculo de un pez saludable o de un pescado recién capturado es estéril, debido a que el sistema inmunológico del pez previene el crecimiento de bacterias en el músculo (Figura 5.8a). Cuando el pez muere, el sistema inmunológico colapsa y las bacterias proliferan libremente. En la superficie de la piel, las bacterias colonizan en una amplia extensión la base de las escamas. Durante el almacenamiento, las bacterias invaden el músculo penetrando entre las fibras musculares. Murray y Shewan (1979), encontraron que sólo un número muy limitado de bacterias invade el músculo durante el almacenamiento en hielo. Ruskol y Bendsen (1992) mostraron mediante exámenes microscópicos que las bacterias pueden ser detectadas en el músculo cuando el número de microorganismos en la superficie de la piel incremente por encima de las 106 ufc/cm2 (Figura 5.6 b). Este resultado fue observado tanto en el almacenamiento en hielo como en ambiente refrigerado. No se encontró diferencia entre los patrones invasivos de las bacterias específicas del deterioro (por ejemplo S. putrefaciens) y las bacterias no específicas del deterioro.
Dado que sólo un número limitado de microorganismos realmente invade el músculo y el crecimiento microbiano se lleva a cabo principalmente en la superficie, el deterioro es probablemente una consecuencia de la difusión de enzimas bacterianas hacia el interior del músculo y de la difusión externa de nutrientes.
El pescado se deteriora a velocidades muy diferentes (véase también Sección 6.5) y se ha propuesto como explicación las diferencias en las propiedades de la superficie del pescado. Las pieles de los peces tienen texturas muy diferentes. Así, el merlán (Merlangius merlangus) y el bacalao (Gadus morhua) que tienen una cubierta muy frágil se deterioran rápidamente en comparación con algunos peces planos como la solla, que posee una dermis y una epidermis robusta. Además, este último grupo cuenta con una gruesa cubierta de mucus, que contiene algunos compuestos antibacterianos, como anticuerpos, complementos y enzimas bacteriolíticas (Murray y Fletcher, 1976; Hjelmland et al., 1983).
Figura 5.8 Sección histológica de: (a) bacalao recién capturado
Cambios en la microflora durante el almacenamiento y deterioro/Organismos específicos del deterioro
Las bacterias presentes en pescados capturados en aguas templadas, entran en fase exponencial de crecimiento casi inmediatamente después de la muerte del pez. Esto también ocurre cuando el pescado es colocado en hielo, probablemente porque la microflora se encuentra adaptada a las temperaturas de enfriamiento. Durante el almacenamiento en hielo, la población bacteriana se duplica en aproximadamente 1 día y después de 2 o 3 semanas alcanza unas 108 - 109 ufc, por gramo de músculo o cm de piel. Durante el almacenamiento a temperatura ambiente, se alcanza un nivel ligeramente inferior a las 10 - 108 ufc/g en 24 horas. Las bacterias presentes en pescados provenientes de aguas tropicales generalmente atraviesan por una fase de latencia de 1 a 2 semanas, cuando el pescado se almacena en hielo, y posteriormente se inicia el crecimiento exponencial. Durante el deterioro, el nivel de bacterias en pescados de aguas tropicales es similar al nivel encontrado en especies de aguas templadas (Gram, 1990; Gram et al, 1990).
Si el pescado en hielo es almacenado en condiciones de anaerobiosis o en una atmósfera de CO2, el número normal de las bacterias psicrotrófas, como la S. putrefaciens y Pseudomonas, es generalmente mucho menor (106 - 107 ufc/g) que en pescado almacenado en condiciones de aerobiosis. Sin embargo, el nivel de bacterias con carácter psicrófilo como P. phosphoreum alcanza las 107 - 108 ufc/g cuando el pescado está deteriorado (Dalgaard et al., 1993).
La composición de la microflora también cambia dramáticamente durante el almacenamiento. De esta forma, después de 1 - 2 semanas de almacenamiento aeróbico en hielo, la flora está constituida casi exclusivamente por Pseudomonas spp. y S. putrefaciens. Esto, se cree, es debido a su relativo corto tiempo de generación a temperaturas de enfriamiento (Morita, 1975; Devaraju y Setty, 1985), este hecho ha sido confirmado por numerosos estudios llevados a cabo en peces de aguas tropicales y de aguas templadas. A temperatura ambiente (25 °C), la microflora en el punto de deterioro está dominada por Vibrionáceas mesofílicas y, particularmente si el pescado proviene de aguas contaminadas, por Enterobacteriáceas.
Debe efectuarse una clara distinción entre los términos flora del deterioro y bacterias del deterioro, dado que el primero describe meramente las bacterias presente en el pescado cuando está deteriorado, mientras que el último se refiere al grupo específico que produce olores y sabores desagradables asociados con el deterioro. Una gran parte de las bacterias presentes en el pescado deteriorado no desempeñan ningún papel en lo absoluto en el deterioro (Figura 5.9). Cada producto pesquero posee sus propias bacterias específicas del deterioro y es el número de estas bacterias, y no el número total de microorganismos, lo que guarda relación con la duración en almacén del producto. En la Figura 5.10, se muestra que el tiempo de vida útil remanente del bacalao en hielo puede ser pronosticado mediante el tiempo de detección conductométrico (en caldo de OTMA), el cual se correlaciona inversamente con el número de puentes de hidrógeno de sulfuro producidos por la acción bacteriana.
No es una tarea fácil determinar, entre las bacterias aisladas del pescado deteriorado, las verdaderas responsables del deterioro, pues se requieren extensos estudios sensoriales, microbiológicos y químicos. En primer lugar deben ser estudiados y cuantificados los cambios sensoriales, microbiológicos y químicos que ocurren durante el almacenamiento, incluyendo la determinación del nivel de un determinado componente químico que se correlacione con deterioro (indicador químico de deterioro). En segundo lugar, se aíslan las bacterias presentes al momento del rechazo sensorial. Poblaciones de bacterias puras y mezcladas se inoculan en sustratos estériles de pescado a fin de evaluar su potencial de deterioro, es decir, su habilidad para producir cambios sensoriales (olores desagradables) y químicos típicos del producto deteriorado. Finalmente, las cepas seleccionadas son examinadas para evaluar su actividad de deterioro, es decir, si su tasa de crecimiento y su producción cualitativa y cuantitativa de olores desagradables son similares a las mediciones en el producto deteriorado (Dalgaard, 1993).
Figura 5.9 Cambios en el recuento total y en las bacterias específicas del deterioro durante el almacenamiento (modificado según Dalgaard (1993))
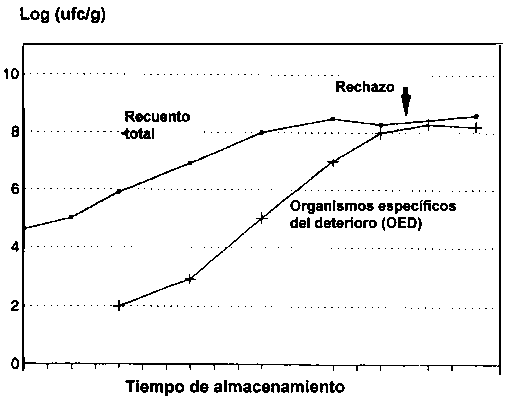
El último paso es particularmente importante, debido a que algunas bacterias pueden producir los compuestos químicos asociados con el deterioro pero son incapaces de hacerlo en cantidades significativas y, por lo tanto, no constituyen bacterias específicas del deterioro. Durante el almacenamiento aeróbico se requieren niveles de 108 - 109 ufc/g de bacterias específicas del deterioro para ocasionar el deterioro. El deterioro del pescado empacado se observa a niveles muy por debajo de 107 ufc de P. phosphoreum por gramo. Este nivel relativamente bajo es debido probablemente al gran tamaño (5 m m) de la bacteria, lo cual origina un mayor rendimiento de por ejemplo la TMA por célula (Dalgaard, 1993).
El potencial y la actividad de deterioro pueden ser medidos en algunos sustratos estériles de pescado como: extracto crudo de pescado (Lerke et al., 1963), extracto de pescado esterilizado por calor (Castell y Greenough, 1957; Gram et al., 1987; Dalgaard, 1993) o en bloques de músculo de pescado estériles (Herbert et al., 1971). Este último es el más complicado pero también es el que proporciona resultados comparables al producto. Si se escoge cualquiera de los extractos del pescado, es importante que la tasa de crecimiento de la bacteria del deterioro en el sistema modelo sea igual a la tasa de crecimiento en el producto.
También puede emplearse una prueba cualitativa para medir la habilidad de la bacteria en producir H2S o reducir OTMA, cuando la flora del deterioro es tamizada en la búsqueda de las bacterias potenciales del deterioro. Un medio donde la reducción del OTMA a TMA puede observarse como el cambio de color de un indicador redox, y la formación de H2S es evidente debido al precipitado negro de FeS desarrollado para este propósito (Gram et al., 1987).
Shewanella putrefaciens ha sido identificada como la bacteria específica del deterioro del pescado de aguas templadas almacenado aeróbicamente en hielo. Si este producto se empaca al vacío, P. phosphoreum participa en el deterioro y pasa a ser la bacteria específica del deterioro del pescado empacado en presencia de CO2 (véase Sección 6.3). La flora del deterioro, del pescado tropical de mar almacenado en hielo, está compuesta casi exclusivamente de Pseudomonas spp. y S. putrefaciens. Algunas Pseudomonas spp. son específicas del deterioro del pescado tropical de agua dulce almacenado en hielo (Lima dos Santos, 1978; Gram et al., 1990) y conjuntamente con S. putrefaciens, son también las causantes del deterioro del pescado marino tropical almacenado en hielo (Gillespie y MacRae, 1975; Gram, 1990).
A temperatura ambiente, las aeromonas móviles son específicas del deterioro del pescado de agua dulce, almacenado aeróbicamente (Gorzyka y Pek Poh Len, 1985; Gram et al., 1990). Barile et al. (1985), demostraron que una gran proporción de la flora en caballa almacenada a temperatura ambiente, estaba constituida por S. putrefaciens, indicando que esta bacteria quizá participa también en el deterioro.
El Cuadro 5.5 proporciona un panorama de las bacterias del deterioro específicas de los productos pesqueros frescos almacenados en hielo y temperatura ambiente.
Cuadro 5.5 Flora dominante y bacterias específicas del deterioro, durante el deterioro de pescado blanco fresco (bacalao) (Huss, 1994)
|
Temperatura de almacenamiento |
Atmósfera de envasado |
Microflora dominante |
Organismos específicos del deterioro (OED) |
Referencias |
|
0°C |
Aeróbica |
Bacilos Gram negativos psicrotróficos, no fermentativos (Pseudomonas spp., S. putrefaciens, Moraxella, Acinetobacter) |
S. putrefaciens Pseudomonas3 |
2, 3, 4, 9 |
|
Vacío |
Bacilos Gram negativos, psicrotróficos o con carácter psicrófilo (S. putrefaciens, Photobacterium) |
S. putrefaciens P. phosphoreum |
1,9 | |
|
EAM¹ |
Bacilos Gram negativos fermentativos con carácter psicrófilo (Photobacterium) Bacilos Gram negativos no fermentativos psicrotróficos (1-10% de la flora: Pseudomonas, S. putrefaciens) Bacilos Gram positivos (BAL2) |
P. phosphoreum |
1,7 | |
|
5°C |
Aeróbica |
Bacilos Gram negativos psicrotróficos (Vibrionáceas, S. putrefaciens) |
Aeromonas spp. S. putrefaciens |
10 |
|
Vacío |
Bacilos Gram negativos psicrotróficos (Vibrionáceas, S. putrefaciens) |
Aeromonas spp. S. putrefaciens |
10 | |
|
EAM |
Bacilos Gram negativos psicrotróficos (Vibrionáceas) |
Aeromonas spp. |
6 | |
|
20 - 30 °C |
Aeróbica |
Bacilos Gram negativos mesófilos fermentativos (Vibrionáceas, Enterobacteriáceas) |
Aeromonas spp. móvil (A. Hydrophila) |
2, 4, 5, 8 |
1) Envasado en atmósfera modificada (que contiene CO2)
2) BAL =
Bacterias acidolácticas
3) En el pescado capturado en aguas tropicales o en
agua dulce suele predominar el deterioro causado por Pseudomonas spp.
Referencias: 1) Barile et al. (1985); 2) Dalgaard et al. (1993); 3) Donald y Gibson (1992); 4) Gorczyca y Pek Poh Len (1985); 5) Gram el al., (1987); 6) Gram et al., (1990); 7) Gram y Dalgaard (comunicación personal); 8) Jorgensen y Huss (1989); 9) Lima dos Santos (1978); 10) van Spreekens (1977)
Cambios bioquímicos inducidos por el crecimiento bacteriano durante el almacenamiento y el deterioro
Al comparar los compuestos químicos desarrollados durante el deterioro natural del pescado y el pescado estéril, se demuestra que la mayoría de los componentes volátiles son producidos por bacterias (Shewan, 1962) según se observa en la Figura 5.11. Estos incluyen trimetilamina, compuestos sulfurosos volátiles, aldehídos, cetonas, ésteres, hipoxantina, así como también otros compuestos de bajo peso molecular.
Los sustratos para la producción de volátiles son los carbohidratos (como el lactado y la ribosa), los nucleótidos (como la inosina monofosfato y la inosina) y otras moléculas de nitrógeno no proteico (NNP). Los aminoácidos son sustratos particularmente importantes para la formación de sulfitos y amoniaco.
Los microorganismos obtienen mucha más energía de la oxidación aeróbica que de la fermentación anaeróbica; así, la completa oxidación de 1 mol de glucosa (u otra hexosa) vía ciclo de Krebs rinde 6 moles de CO2 y 36 moles de ATP. Por el contrario, la fermentación de 1 mol de glucosa rinde sólo 2 moles de ATP y dos moles de ácido láctico. El crecimiento aeróbico inicial en pescado es dominado por bacterias que utilizan carbohidratos como sustrato y oxígeno como aceptor terminal de electrones, con la concomitante producción de CO2 y H2O.
Reducción del Oxido de Trimetilamina (OTMA)
El crecimiento de bacterias consumidoras de oxígeno ocasiona la formación de nichos anaeróbicos o microaerofílicos en el pescado. Esto sin embargo no necesariamente favorece el crecimiento de bacterias anaeróbicas. Algunas de las bacterias presentes en el pescado son capaces de llevar a cabo respiración (con la ventaja del ATP) empleando otras moléculas como receptor final del electrón. Es típico de muchas bacterias específicas del deterioro del pescado emplear el OTMA como aceptor terminal de electrones durante la respiración anaeróbica. El componente reducido, la TMA; uno de los compuestos dominantes del pescado deteriorado, tiene el olor típico del pescado. El nivel de TMA encontrado en pescado fresco rechazado por un panel sensorial varía dependiendo de la especie de pescado, pero generalmente se encuentra alrededor de los 10-15 mg TMA-N/100 g en pescado almacenado aeróbicamente y en un nivel de 30 mg TMA-N/100 g en bacalao empacado (Dalgaard et al., 1993).
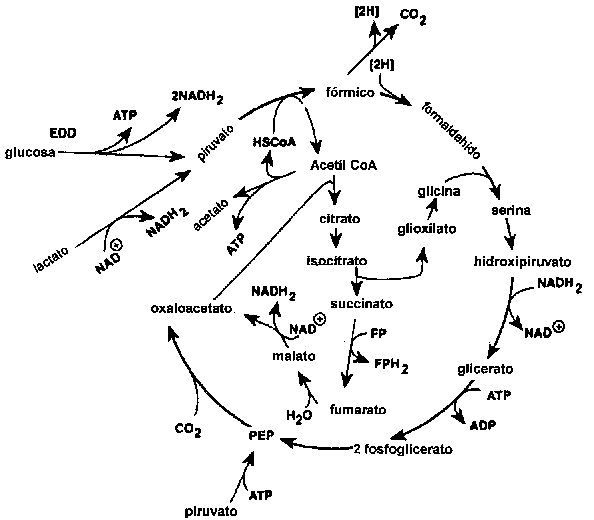
La reducción del OTMA está generalmente asociada con géneros de bacterias típicos del ambiente marino (Alteromonas, Photobacterium, Vibrio y S. putrefaciens), pero también es llevada a cabo por Aeromonas y bacterias intestinales de las Enterobacteriáceas. La reducción del OTMA ha sido estudiada en bacterias fermentativas, anaerobias facultativas, como E. coli (Sakaguchi et al., 1980) y Proteus spp. (Stenberg et al., 1982) como también en la bacteria no fermentativa S. putrefaciens (Easter et al., 1983; Ringo et al., 1984). Durante el crecimiento aeróbico, S. putrefaciens emplea el ciclo de Krebs para producir los electrones que posteriormente son canalizados a través de la cadena respiratoria. Ringo et al. (1984) proponen que durante la respiración anaeróbica S. putrefaciens también utiliza todo el ciclo de Krebs (Figura 5.12), mientras recientemente se ha demostrado que en la respiración anaeróbica de S. putrefaciens, sólo utiliza una parte del ciclo de Krebs (Figura 5.13) y los electrones son generados también por otra ruta metabólica, denominada la ruta de la serina (Scott y Nealson, 1994). S. putrefaciens puede emplear una variedad de fuentes de carbono como sustrato en su respiración anaeróbica dependiente de OTMA, incluyendo formato y lactato. Compuestos como acetato y succinato empleados en la respiración del oxígeno no pueden ser empleados cuando el OTMA es el aceptor terminal de electrones (DiChristina y DeLong, 1994), por el contrario, el acetato es uno de. los productos del la reducción anaeróbica del OTMA (Ringo et al., 1984; Scott y Nealson, 1994).
Figura 5.12 Reducción anaeróbica del OTMA por S. putrefaciens (anteriormente Alteromonas) según propuesta de Ringo et al. (1984)
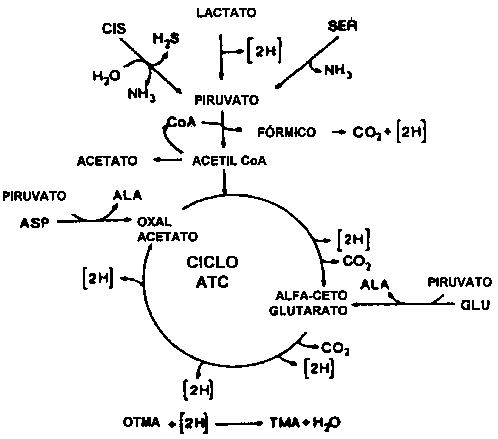
Figura 5.13 Ruta del carbón durante la anaerobiosis propuesta para S. putrefaciens (Scott y Nealson, 1994)
Por el contrario, los azúcares y el lactato son los principales sustratos generadores de electrones cuando el OTMA es reducido por Proteus spp. La reducción está acompañada por la producción de acetato como producto principal (Kjosbakken y Larsen, 1974).
El OTMA es un compuesto típico de los peces marinos, según se mencionó en la Sección 4.4, y recientemente ha sido reportado que también algunos peces de agua dulce contienen altas cantidades de OTMA (Anthoni et al., 1990). Sin embargo, la TMA no es necesariamente un compuesto característico durante el deterioro de este tipo de pescado porque el deterioro es debido a Pseudomonas spp. (Gram et al., 1990).
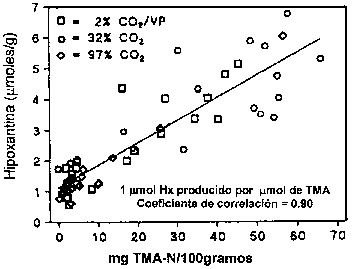
En muchas especies de pescado el desarrollo de la TMA es paralelo a la producción de hipoxantina. La hipoxantina, según lo descrito en la Sección 5.2, puede ser formada por la descomposición autolítica de nucleótidos, pero también puede ser formada por bacterias; la tasa de formación por la acción bacteriana es mayor que por autólisis. Tanto Jorgensen et al. (1988) como Dalgaard (1993) demostraron una correlación linear entre el contenido de TMA e hipoxantina durante el almacenamiento en hielo de bacalao empacado (Figura 5.14). Algunas de las bacterias del deterioro producen hipoxantina a partir de inosina o inosina monofosfato, incluyendo Pseudomonas spp. (Surette et al., 1988), S. putrefaciens (van Spreekens, 1977; Jorgensen y Huss, 1989; Gram, 1989) y P. phosphoreum (van Spreekens, 1977).
En el bacalao y en otros gádidos, hasta que ocurre el deterioro, la TMA constituye la mayor parte de las denominadas bases volátiles totales; BVT (también conocidas como nitrógeno volátil total, NVT). Sin embargo, en el pescado deteriorado el suministro de OTMA decae, la TMA alcanza su máximo nivel y los niveles de NVT continúan incrementando debido a la formación de NH3 y otras aminas volátiles. En las primeras semanas del almacenamiento en hielo también se forma un poco de amoniaco debido a la autólisis. En algunos pescados que no contienen OTMA, o en los cuales el deterioro es debido a una flora no reductora de OTMA, se observa un leve incremento en las BVT durante el almacenamiento, probablemente como resultado de la desaminación de aminoácidos.
Figura 5.14 Relación entre el contenido de TMA e Hx durante el almacenamiento de bacalao empacado en hielo (Dalgaard et al., 1993)
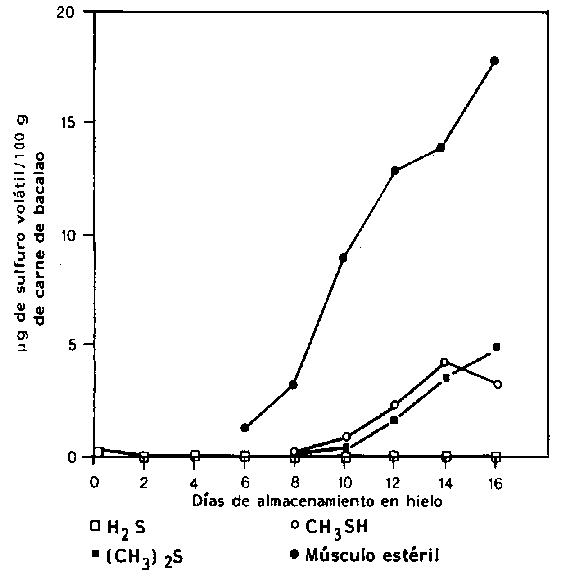
Los compuestos sulfurados volátiles son componentes típicos del pescado deteriorado y la mayoría de las bacterias identificadas como bacterias específicas del deterioro producen uno o algunos sulfuros volátiles. S putrefaciens y algunas Vibrionaceae producen H2S a partir del aminoácido sulfurado 1-cisteína (Stenstroem y Molin, 1990; Gram et al., 1987). Por el contrario, ni Pseudomonas o P. phosphoreum producen cantidades significativas de H2S. De esta forma el sulfuro de hidrógeno, compuesto típico del deterioro del bacalao almacenado aeróbicamente en hielo, no se produce durante el deterioro del pescado empacado en CO2 (Dalgaard et al., 1993). El metilmercaptano (CH3SH) y el dimetilsulfuro ((CH3)2S) son formados a partir del otro aminoácido sulfurado, la metionina. La taurina, que también contiene sulfuro, se presenta como aminoácido libre en muy altas concentraciones en el músculo del pescado. Este aminoácido desaparece del músculo del pescado durante el almacenamiento (Figura 5.11), pero debido más al goteo que al ataque bacteriano (Herbert y Shewan, 1975). En la Figura 5.15 se muestra la formación de compuestos en el bacalao deteriorado naturalmente, comparado con el músculo estéril.
Los compuestos sulfurados volátiles tienen un olor muy desagradable y pueden ser detectados hasta en niveles de ppb, incluso estas mínimas cantidades tienen un efecto considerable en la calidad.
Ringo et al. (1984) han demostrado que la cisteína es utilizada como sustrato en el ciclo de Krebs cuando los electrones son transferidos al OTMA, de este modo la formación de H2S y TMA son hasta cierto punto reacciones vinculadas (Figura 5.12).
Figura 5.15 Producción de H2S, CH3SH y (CH3)2S, en filetes de bacalao deteriorados naturalmente y en bloques de músculo estéril (Herbert y Shewan, 1976)
Contrario al deterioro en hielo por S. putrefaciens y al deterioro a temperatura ambiente por Vibrionaceae dominados por la producción de H2S y TMA, el deterioro causado por Pseudomonas spp, está caracterizado por la ausencia de estos compuestos (Gram et al., 1989, Gram et al., 1990). El deterioro del pescado almacenado en hielo por Pseudomonas genera olores y sabores desagradables arrutados, a podrido y a sulfuro. Las Pseudomonas spp. producen un número de compuestos volátiles, como aldehídos, cetonas, ésteres y sulfuros (Edwards et al., 1987; Miller et al., 1973a, 1973b). Sin embargo, se desconoce el compuesto específico responsable de los típicos olores desagradables (Cuadro 5.6). Los olores afrutados desagradables producidos por Pseudomonas fragi se originan a partir de los monoaminoácidos y los aminoácidos monocarboxílicos.
Cuadro 5.6 Compuestos típicos del deterioro, producidos durante el deterioro del pescado fresco almacenado aeróbicamente, o empacado en hielo o a temperatura ambiente
|
Organismo específico del deterioro |
Compuesto típico del deterioro |
|
Shewanella putrefaciens |
TMA, H2S, CH3SH, (CH3)2S y Hx |
|
Photobacterium phosphoreum |
TMA, Hx |
|
Pseudomonas spp. |
Cetonas, aldehídos, ésteres, sulfuros no-H2S |
|
Vibrionaceae |
TMA, H2S |
|
Anaeróbicos deteriorativos |
NH3, ácidos: acético, butírico y propiónico |
Según se mencionó anteriormente, el nivel de las BVT continúa incrementando incluso después que la TMA ha alcanzado su máximo. Lo anterior es debido a la proteólisis que se inicia cuando algunos de los aminoácidos libres han sido utilizados. Lerke et al. (1967) separaron extracto de pescado en fracciones proteicas y no proteicas, e inocularon bacterias del deterioro en cada fracción y en el extracto total. La fracción no proteica del extracto de pescado se deterioró como todo el extracto, mientras que en la fracción proteica del extracto sólo se detectaron leves olores desagradables. Aunque, algunos autores han empleado el número de bacterias proteolíticas como un indicador del deterioro, se debe concluir que el volumen de la fracción proteica es de menor importancia en el deterioro del pescado fresco.
Algunos de los compuestos típicos formados por las bacterias durante el deterioro del pescado se muestran en el Cuadro 5.7, conjuntamente con los sustratos empleados para su formación.
Cuadro 5.7 Sustratos y compuestos, de olores y sabores desagradables, producidos por las bacterias durante el deterioro del pescado
|
Sustrato |
Compuestos producidos por la acción bacteriana |
|
OTMA |
TMA |
|
cisteína |
HsS |
|
metionina |
CH3SH, (CH3)2S |
|
carbohidratos y lactato |
acetato, CO2, H2O |
|
inosina, IMP |
hipoxantina |
|
aminoácidos (glicina, serina, leucina) |
ésteres, cetonas, aldehídos |
|
aminoácidos, urea |
NH3 |
La formación de TMA está acompañada por la formación de amoniaco durante el almacenamiento anóxico del arenque y la caballa (Haaland y Njaa, 1988). El almacenamiento anaeróbico prolongado del pescado ocasiona una vigorosa producción de NH3, debido a la degradación posterior de aminoácidos y a la acumulación de ácidos grasos como los ácidos acético, butírico y propiónico. Se determinó que los más fuertes productores de NH3 son anaerobios obligados pertenecientes a la familia Bacteroidaceae género Fusobacterium (Kjosbakken y Larsen, 1974; Storroe et al., 1975, 1977). Estos organismos sólo crecen en el extracto de pescado deteriorado y tienen muy poca o ninguna actividad proteolítica, por lo cual emplean proteínas ya hidrolizadas.
Durante el almacenamiento en hielo del pescado graso fresco, los cambios en la fracción lipídica son causados casi exclusivamente por la acción química, por ejemplo: la oxidación, por cuanto el ataque bacteriano en la fracción lipídica contribuye muy poco al perfil de deterioro. Durante el almacenamiento del pescado ligeramente preservado, la hidrólisis lipídica causada por bacterias puede ser parte del perfil de deterioro.
En los lípidos del pescado ocurren dos reacciones diferentes, de importancia en el deterioro de la calidad:
· oxidación
· hidrólisis
Ellas dan como resultado la producción de una serie de sustancias, de las cuales algunas tienen sabores y olores desagradables (rancio). Algunas pueden también contribuir a los cambios de textura mediante uniones covalentes a las proteínas musculares. Las reacciones pueden ser no enzimáticas o catalizadas por enzimas: microbianas, intracelulares o digestivas del mismo pescado. Por lo tanto, el significado relativo de estas reacciones depende principalmente de la especie de pescado y de la temperatura de almacenamiento.
Los pescados grasos son, por su puesto, particularmente susceptibles a la degradación lipídica, la cual puede ocasionar severos problemas en la calidad, incluso durante el almacenamiento a temperaturas bajo cero.
Oxidación
La gran cantidad de ácidos grasos poliinsaturados presente en los lípidos del pescado (véase sección 4.2) les hace altamente susceptibles a la oxidación mediante un mecanismo autocatalítico (Figura 5.16). El proceso es iniciado, según se describe más adelante, mediante la escisión de un átomo de hidrógeno del átomo de carbono central de la estructura pentahédrica presente en la mayoría de las acilcadenas de los ácidos grasos con más de un doble enlace:
-CH=CH-CH2-CH=CH ® -CH=CH-CH-CH-CH- +H·
Contrario a la molécula nativa, el radical lipídico (L·) reacciona muy rápidamente con el oxígeno atmosférico formando un radical peróxido (LOO·), el cual puede nuevamente escindir un hidrógeno de otra acilcadena produciendo un hidroperóxido (LOOH) y un nuevo radical L·. Esta propagación continúa hasta que uno de los radicales es removido mediante reacción con otro radical o con un antioxidante (AH) del cual resulta un radical (A·) mucho menos reactivo. Los hidroperóxidos, producidos en cantidades relativamente grandes durante la propagación, son insípidos y, por lo tanto, quizá no es una sorpresa que el ampliamente usado "valor de peróxido" (Sección 8.2) generalmente guarda escasa correlación con las propiedades sensoriales.
Figura 5.16 Autooxidación de un lípido poliinsaturado
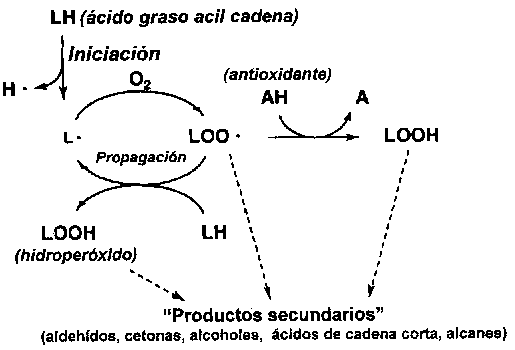
Los hidroperóxidos continúan dividiéndose, catalizados por iones de metales pesados, hasta la formación de cadenas carbonadas más cortas, productos secundarios de la autooxidación. Estos productos secundarios -principalmente aldehídos, cetonas, alcoholes, pequeños ácidos carboxílicos y alcanes- originan un extenso espectro de olores y en algunos casos decoloración amarillenta. Algunos de los aldehídos pueden ser determinados como "sustancias reactivas al ácido tiobarbitúrico" (Sección 8.2).
Los iones metálicos son de gran importancia en el primer paso de la autooxidación de los lípidos - el proceso de iniciación - como catalizadores de la formación de especies reactivas al oxígeno, como por ejemplo: el radical hidróxilo (OH·). Este radical reacciona inmediatamente con los lípidos o cualquier otra molécula en el lugar donde ha sido generado. La alta reactividad quizá explique el hecho de que los ácidos grasos libres sean más susceptibles a la oxidación que los correspondientes ácidos grasos no libres, debido a que la cantidad de hierro en la fase acuosa es probablemente mayor que la cantidad enlazada a la superficie de las membranas celulares y a las gotas de lípidos.
Los hidroperóxidos de los ácidos grasos pueden también ser formados enzimáticamente, catalizados por la enzima lipoxigenasa, la cual está presente en los diferentes tejidos del pescado en cantidades variables. La enzima es inestable y probablemente tiene importancia en la oxidación de los lípidos sólo en el pescado fresco. La cocción o las operaciones de congelado/descongelado destruyen efectivamente la actividad de la enzima.
La célula viva posee algunos mecanismos de protección dirigidos contra los productos de la oxidación lipídica. Existe una enzima, la glutatión peroxidasa, que reduce los hidroperóxidos en las membranas celulares al correspondiente compuesto hidroxílico. Esta reacción requiere un suministro de glutatión reducido y por lo tanto cesa cuando el pez muere y la sustancia se agota en la célula. Las membranas también contienen un compuesto fenólico, a -tocoferol (Vitamina E), el cual es considerado como el más importante antioxidante natural. El tocoferol puede donar átomos de hidrógeno a los radicales L· o LOO · funcionando como la molécula AH en la Figura 5.16. Se asume que el radical tocoferil resultante reacciona con el ácido ascórbico (Vitamina C) en la interfase lípido/agua regenerándose la molécula de tocoferol. Otros compuestos, por ejemplo los carotenoides, pueden también funcionar como antioxidantes. El humo de la madera contiene fenoles los cuales pueden penetrar la superficie del pescado durante el ahumado y proporcionar de esta forma alguna protección contra la oxidación lipídica.
Hidrólisis
Durante el almacenamiento, aparece una cantidad considerable de ácidos grasos libres (AGL) (Figura 5.17). El fenómeno es más profundo en el pescado no eviscerado que en el eviscerado, probablemente por las enzimas digestivas. Los triglicéridos presentes en los depósitos de grasas son escindidos por la trigliceril lipasa (TL in la Figura 5.18) originada del tracto digestivo o excretada por ciertos microorganismos. Las lipasas celulares pueden también desempeñar un papel menor.
Figura 5.17 Desarrollo de ácidos grasos libres en arenque almacenado a diferentes temperaturas (Laboratorio Tecnológico, Ministerio de Pesca de Dinamarca, Reporte Anual, 1971)
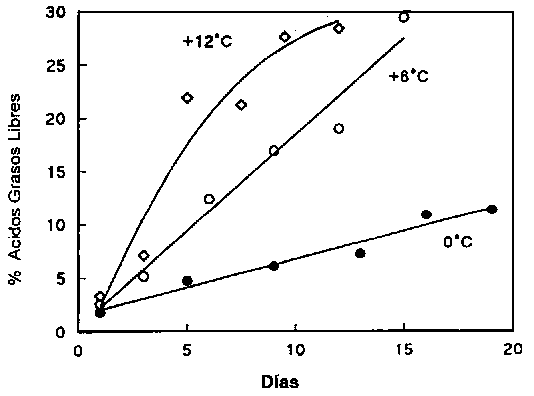
Figura 5.18 Reacciones hidrolíticas primarias de triglicéridos y fosfolípidos. Enzimas: PL1 y PL2, fosfolipasas; TL, trigliceril lipasa
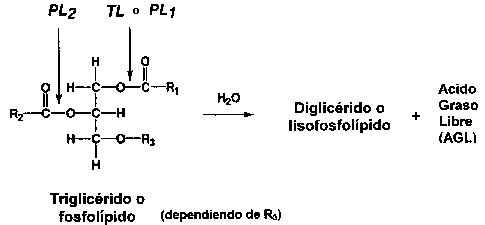
En el pescado magro, por ejemplo: el bacalao del Atlántico, la producción de ácidos grasos libres ocurre incluso a bajas temperaturas. Se cree que las enzimas responsables son fosfolipasas celulares - particularmente la fosfolipasa A2. (PL2 en la Figura 5.18) - a pesar de que aún no ha sido establecida plenamente una correlación entre la actividad de estas enzimas y la tasa de aparición de los ácidos grasos libres. Los ácidos grasos que están unidos a fosfolípidos en el átomo de carbono 2 del glicerol, son principalmente del tipo poliinsaturados; en tal sentido, la hidrólisis generalmente también conduce a incrementar la oxidación. Además, los ácidos grasos por sí mismos pueden causar un sabor jabonoso.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}